Иксодовые клещи — паразиты и переносчики инфекций
Покровы иксодовых клещей
Тело иксодовых клещей покрыто плотными покровами, выполняющими роль наружного скелета и благодаря высокой водонепроницаемости препятствующими испарению воды. Строение покровов иксодид подробно описано для Boophilus (Beadle, 1974), H. asiaticum (Атлас..., 1979), I. persulcatus (Таежный клещ..., 1985) и H. dromedarii (El Shoura, 1988a). У всех этих видов строение покровов было сходным. В состав покровов входит слой гиподермальных клеток (гиподерма) и выделяемая ими кутикула, которая имеет сложное строение. Внутренняя часть кутикулы, образующая основную ее массу и лежащая непосредственно над гиподермой, носит название прокутикулы.
Прокутикула построена из связанных между собой молекул протеинов и хитина. Биохимическими исследованиями установлено, что для прокутикулы иксодовых клещей характерны относительно низкое содержание хитина (около 4 %) и своеобразный аминокислотный состав белков. Эти особенности кутикулы, возможно, обусловливают исключительную способность некоторых участков покровов к растяжению. В зависимости от степени склеротизации и способности к растяжению различают три вида прокутикулы: эндокутикулу, мезокутикулу и экзокутикулу. Эндокутикула — несклеротизованная растяжимая прокутикула; экзокутикула — полностью задубленная; в мезокутикуле процессы склеротизации еще не завершены. В тех случаях, когда в состав кутикулы входят все три вида прокутикулы, эндкутикула располагается над слоем гиподермальных клеток, мезокутикула — над ней, а экзокутикула образует внешний, наиболее удаленный от гиподермы слой.
Из эндокутикулы построена растяжимая часть идиосомы недавно перелинявших клещей. В дальнейшем, на стадии послелиночного доразвития происходит частичная склеротизация прокутикулы в этой части покровов и превращение ее в мезокутикулу. Толщина эндо- и мезокутикулы голодных самок таежного клеща составляет соответственно 3—4 и 15—18 мкм, а у H. asiaticum они вдвое толще. У нимф толщина прокутикулы меньше, чем у взрослых, а у личинок она тоньше, чем у нимф, и не превышает 2—8 мкм. На электронограммах мезокутикула (табл. XIV, 1—3) имеет тонкогранулярную структуру и обладает умеренной электронной плотностью. Она пронизана слегка извилистыми поровыми каналами диаметром 100—200 нм, в просвете которых иногда содержится гранулярный или фиброзный материал. Эндокутикула имеет выраженную ламеллярную структуру. Каждая ламелла состоит из тонких микрофибрилл, образующих сложный рисунок; толщина ламеллы 1.5—2 мкм. Поровые каналы этой части кутикулы имеют дугообразную форму.
Только из экзокутикулы образована кутикула твердых (нерастяжимых) покровов — спинного щитка, вентральных щитков самца, анального клапана, перитрем, гнатосомы, конечностей. Экзокутикула не окрашивается без специальной предварительной обработки и имеет на срезах янтарно-желтый оттенок. На электронограммах она выглядит гомогенной, но несколько более электронно-плотной по сравнению с мезокутикулой.
Поверхность кутикулы таежного клеща образует регулярные складки (табл. IX, 1; X, 1, 2) и, так же как у других наземных членистоногих, покрыта эпикутикулой, которая при исследовании в световом микроскопе выглядит однородной. В отличие от прокутикулы для эпикутикулы характерно отсутствие хитина. Толщина этого слоя, по данным световой микроскопии, составляет 1—4 мкм.
Строение эпикутикулы иксодид одинаково во всех частях покровов независимо от степени их склеротизации. По данным электронной микроскопии, она имеет сложное строение и состоит из четырех слоев, различающихся по толщине и электронной плотности (рис. 4). Самый внешний из слоев эпикутикулы, восковой, имеет толщину около 100—200 нм. Для него характерна умеренная электронная плотность, и иногда в нем выявляется горизонтальная слоистость. Лежащий на нижней границе воскового очень тонкий (2—4 нм) электронно-плотный слой, по-видимому, образован мономолекулярным слоем липидов. Следующий, кутикулиновый, слой отличается от остальных частей эпикутикулы чрезвычайно высокой электронной плотностью, его толщина составляет 15—30 нм. Этот слой первым откладывается при образовании кутикулы во время линьки. Под кутикулиновым слоем располагается протеиновая эпикутикула, близкая по структуре и электронной плотности к прокутикуле, с которой граничит. Толщина этой части эпикутикулы 150—300 нм.
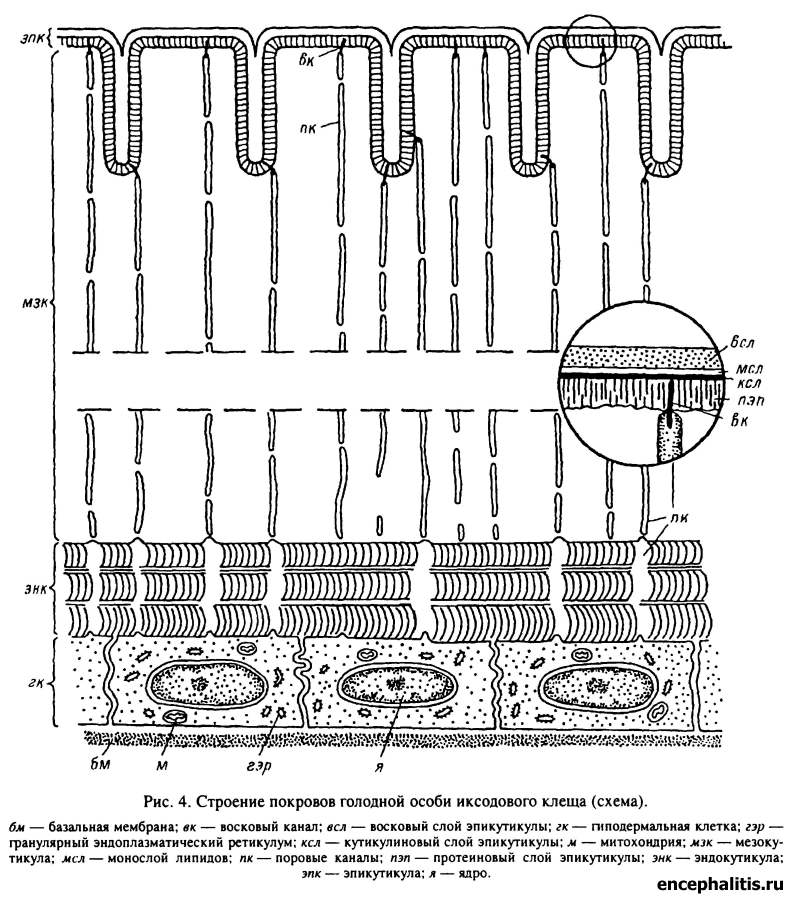
Гиподерма представлена одним слоем клеток. Клетки голодных особей имеют уплощенную форму, их высота составляет от 1—2 мкм у личинок до 5—10 мкм у взрослых. Ультраструктура гиподермальных клеток голодных особей (табл. XIV, 2) свидетельствует об отсутствии в них активных синтетических и секреторных процессов. В цитоплазме слабо развиты элементы гранулярного эндоплазматического ретикулума и содержится лишь незначительное количество свободных рибосом.
Строение покровов значительно изменяется в период многодневного питания, во время которого происходят рост и растяжение кутикулы. В течение большей части периода питания наблюдается увеличение толщины кутикулы, причем отмечен рост как эндокутикулы, так и мезокутикулы.
У самок таежного клеща толщина эндокутикулы увеличивается с 3—4 до 17— 30 мкм, а мезокутикулы — с 15—18 до 30—32 мкм. Одновременно отмечается и возрастание сухой массы кутикулы. В конце периода питания рост кутикулы прекращается и происходит ее растяжение, при этом толщина мезокутикулы снижается до 10—12, а эндокутикулы — до 9—10 мкм. О растяжении кутикулы свидетельствуют увеличение диаметра поровых каналов и изменения в ультраструктуре кутикулярных ламелл. Растяжение сопровождается расправлением борозд и регулярных складок на поверхности растяжимой кутикулы идиосомы.
Рост кутикулы обеспечивается секреторной активностью гиподермы, в строении которой в это время наблюдаются значительные изменения. Высота клеток увеличивается в 3—4 раза по сравнению с периодом голодания и достигает 30— 35 мкм, а форма клеток становится столбчатой. В конце периода питания толщина гиподермального слоя вновь несколько уменьшается, что связано с интенсивным растяжением покровов, происходящим в это время. С началом питания увеличиваются и размеры ядер и ядрышек. В гиподермальном слое часто можно наблюдать делящиеся клетки. Цитоплазма становится базофильной, в ней выявляются РНК и щелочная фосфатаза. Повышается электронная плотность цитоплазмы, в ней появляются многочисленные цистерны гранулярного эндоплазматического ретикулума и комплексы Гольджи, а в апикальных частях клетки, граничащих с кутикулой (табл. XIV, 4), — секреторные пузырьки нескольких видов. На поверхности клеток, обращенной к кутикуле, возрастает количество микроворсинок, по ультраструктуре значительно отличающихся от микроворсинок голодных особей. Они, по-видимому, принимают участие в образовании и ориентации хитиново-протеиновых фибрилл прокутикулы.
Гистологические изменения, происходящие в период линьки, однотипны у всех иксодид. В отличие от других кровососов у иксодид процессы роста покровов и внутренних органов, связанные с питанием, переходят в процессы подготовки к линьке. Рост продолжается и в течение нескольких дней, которые отделяют момент отпадения напитавшегося клеща от начала линочных изменений. В этот период гиподермальные клетки продолжают активно секретировать. Вероятно, одновременно происходит изменение химического состава покровов и усиление их склеротизации, благодаря чему возрастают их твердость и хрупкость.
Перед началом линьки рост кутикулы прекращается. Далее происходит отделение кутикулы от гиподермы-(или аполизис), которое, с одной стороны, представляет следствие митотической активности гиподермы, а с другой — является результатом растворения нижних ламелл эндокутикулы под действием секрета гиподермы. Пространство между гиподермой и кутикулой, возникающее в результате их отделения друг от друга, носит название экзувиальной полости. Ее образование начинается с переднего конца тела. Экзувиальная полость заполнена линочной жидкостью. Содержимое экзувиальной полости в процессе формирования новой кутикулы частично всасывается гиподермальными клетками.
Откладка кутикулы следующей фазы развития начинается с послойного образования эпикутикулы. Первым откладывается кутикулиновый слой. Вслед за кутикулиновым слоем откладывается протеиновая эпикутикула. В то же время начинается формирование кутикулярных складок. Первым шагом к образованию поверхностного рисунка является формирование на апикальной поверхности гиподермы, покрытой двумя слоями эпикутикулы, регулярных углублений. Микроворсинки в промежутках между ними значительно удлиняются. Над концами и вокруг микроворсинок происходит откладка материала прокутикулы. По мере образования прокутикулы внутри складок на месте микроворсинок формируются поровые каналы. В период откладки основной массы прокутикулы микроворсинки укорачиваются до своей обычной длины (около 0.5 мкм), а в образовании поровых каналов принимают участие цитоплазматические выросты на поверхности гиподермальных клеток. Формирование эпикутикулы завершается уже после сбрасывания линочной шкурки, когда на поверхности кутикулы появляется восковой слой. Откладка различных компонентов кутикулы сопровождается активной секреторной деятельностью гиподермы.
В начале периода линьки — во время откладки кутикул и нового слоя и протеиновой эпикутикулы наблюдается скопление в базальных частях гиподермальных клеток больших количеств гликогена. Как показали электронно-микроскопические исследования, скопления гликогена занимают практически всю подъядерную зону клеток, не оставляя места для других видов включений и органоидов. К моменту сбрасывания старой кутикулы количество гликогена значительно уменьшается.
Активные синтетические процессы продолжаются в гиподермальных клетках и после окончания линьки. В период послел и ночного доразвития происходит склеротизация покровов и на этом завершается формирование эпикутикулы.
Помимо обычных гиподермальных клеток в состав покровов иксодовых клещей входят специализированные железистые клетки, связанные с выводным протоком, открывающимся на поверхность кутикулы. Эти образования получили название дермальных желез. Дермальные железы встречаются по всей поверхности растяжимой кутикулы идиосомы, на спинном щитке и на основании гнатосомы. Они открываются на поверхность сложно устроенными кутикулярными протоками, которые у желез I типа иногда называют копьевидными сенсиллами и у желез II типа — стреловидными.
У I. persulcatus обнаружены только дермальные железы I типа. Они состоят из двух железистых клеток и нескольких клеток, образующих проток (рис. 5).
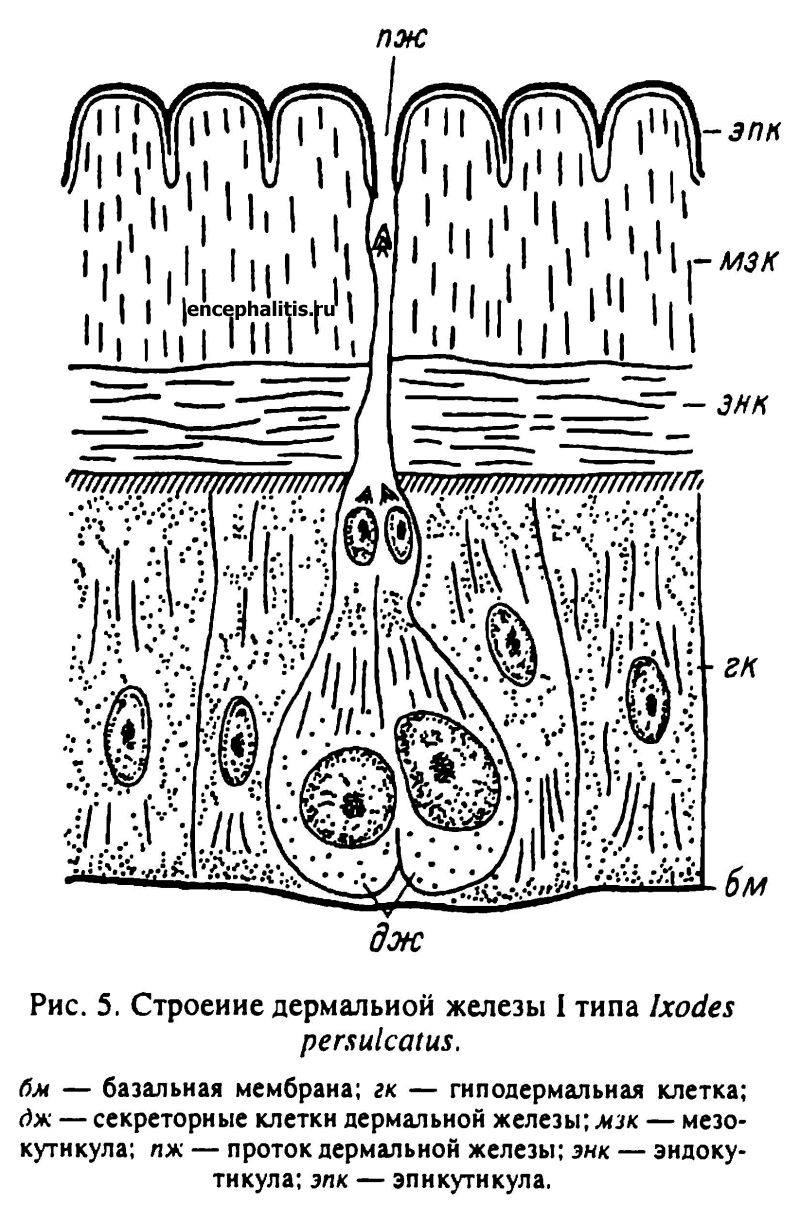
У голодных особей железистые клетки практически неотличимы от окружающих клеток гиподермы. Активизация их секреторной деятельности происходит с началом питания. В это время железистые клетки и их ядра увеличиваются в объеме. Наблюдается усиление базофилии цитоплазмы. В апикальной части клетки обычно появляется крупная вакуоль, которая, по данным электронной микроскопии, представляет обширную инвагинацию поверхностной мембраны железистой клетки, сообщающуюся с протоком железы. Поверхность инвагинации несет многочисленные микроворсинки. В ультраструктуре цитоплазмы также происходят значительные изменения: резко увеличивается количество элементов гранулярного эндоплазматического ретикулума, появляются развитые комплексы Гольджи и многочисленные секреторные вакуоли. Железистые клетки, очевидно, продолжают активно секретировать и у клещей, закончивших питание, однако с началом линочных изменений происходит их дегенерация, хотя ядра железистых клеток сохраняются вплоть до начала откладки эпикутикулы.
У Н. asiaticum (Атлас..., 1979), R. appendiculatus (Walker et al., 1996a, 1996b) и других амблиомин кроме описанных выше желез I типа имеются также более крупные железы II типа. Они также состоят из 2 железистых клеток, которые уже у голодных особей значительно крупнее гиподермальных (рис. 6). С началом кровососания размеры их быстро увеличиваются и достигают у напитавшихся самок 200—250 и у нимф 100 мкм. После отпадения напитавшихся особей размеры желез продолжают некоторое время увеличиваться и достигают 300—400 мкм при диаметре ядер 40—80 мкм. Апикальная часть каждой из железистых клеток заполняется секреторными вакуолями, содержимое которых изливается в обширную полость сообщающуюся с выводным протоком железы. Именно у напитавшихся особей секрет этих желез изливается на поверхность покровов в виде мелких капелек жидкости у отверстий их стреловидных протоков. Затем начинается их быстрая дегенерация, полость спадается и железа исчезает.
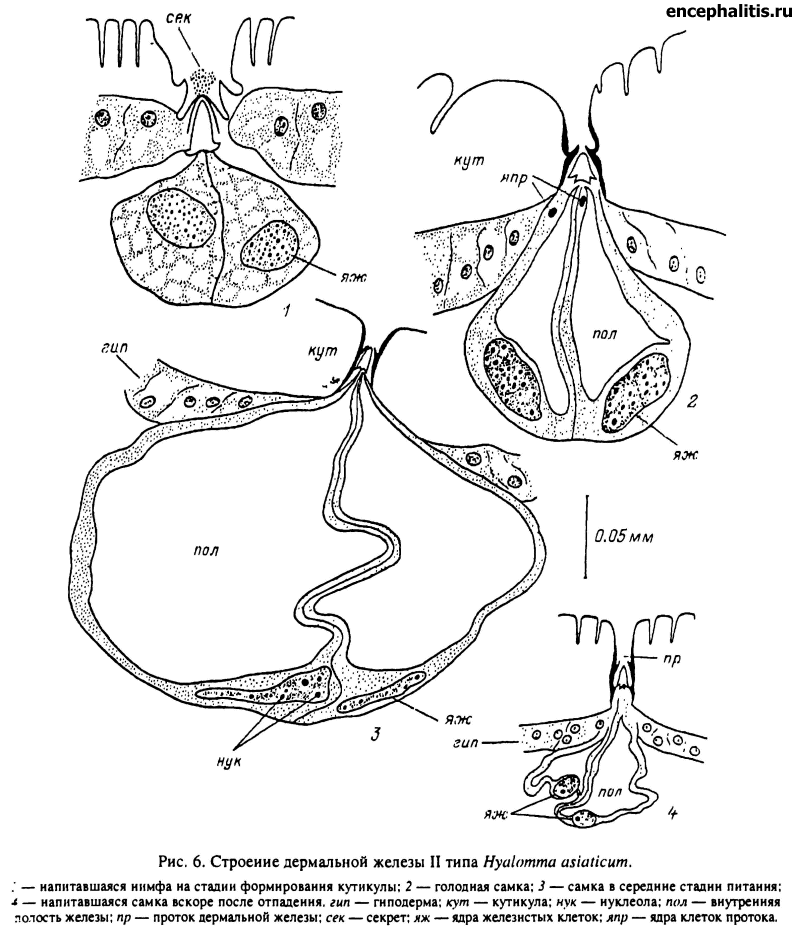
Функции дермальных желез как таежного клеща, так и других иксодовых клещей остаются неясными. Некоторые исследователи (Лагугенко, 1969) описывали их как железисто-чувствительные органы — копьевидные сенсиллы и приписывали секрету функцию смазки, предохраняющей чувствительные волоски. Однако дальнейшие исследования не подтвердили этого предположения. Дермальные железы образуют белковые и липоидные секреты, часть из которых участвует в феромонной коммуникации клещей (Walker et al., 1996b).
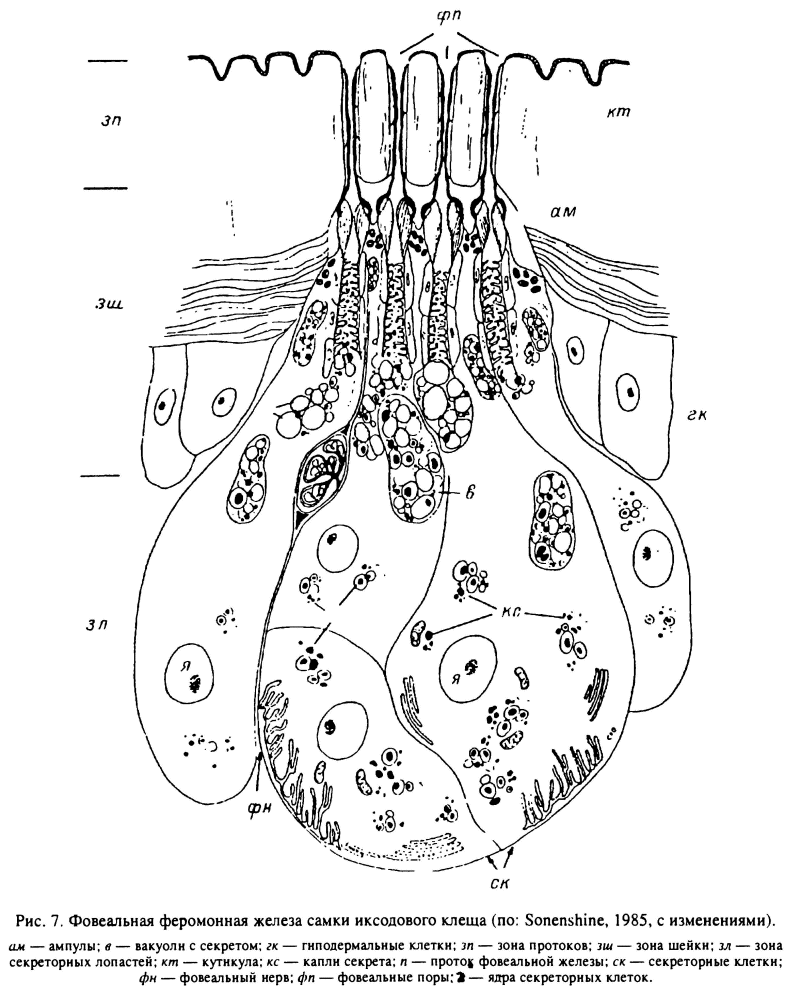
Специализированным типом дермальных желез являются так называемые фовеальные железы, подробно исследованные у H. dromedarii (Sonenshine et al., 1983). На поверхности фовеа открываются выводные протоки нескольких десятков подобных желез. Каждая железа включает клетки протока и 3 железистые клетки. У голодных самок фовеальные железы находятся в неактивном состоянии. С началом питания размеры железистых клеток увеличиваются в несколько раз и цитоплазма их заполняется секреторными включениями (рис. 7). Предполагают, что продуктом их секреции служит летучий половой феромон — 2,6-дихлорфенол, выделяемый через отверстия в кутикуле фовеа питающихся самок.
Особые дермальные железы обнаружены и в тарзальных члениках каждой из 4 пар ног H. asiaticum. Они образованы из многих клеток однослойного эпителия, окружающих мешковидную центральную полость железы. Полость железы открывается наружу выстланным кутикулой протоком. Клетки стенок железы заполнены секреторными гранулами.
Специализированная группа дермальных желез расположена под поровыми полями. Протоки этих желез отрываются на поровых полях, а их секрет участвует в формировании водонепроницаемого покрытия откладываемых яиц (Kakuda et al., 1995а).