Иксодовые клещи — паразиты и переносчики инфекций
Кишечник и особенности пищеварения
Питание каждой фазы таежного клеща представляет собой многодневную стадию развития на теле хозяина (см. главу VI). На этой стадии клещ не только поглощает огромные количества пищи, но и переваривает значительную часть ее. Кишечник клещей на этой стадии многократно увеличивается в объеме и заполняет большую часть идиосомы (рис. 25, 1—3).
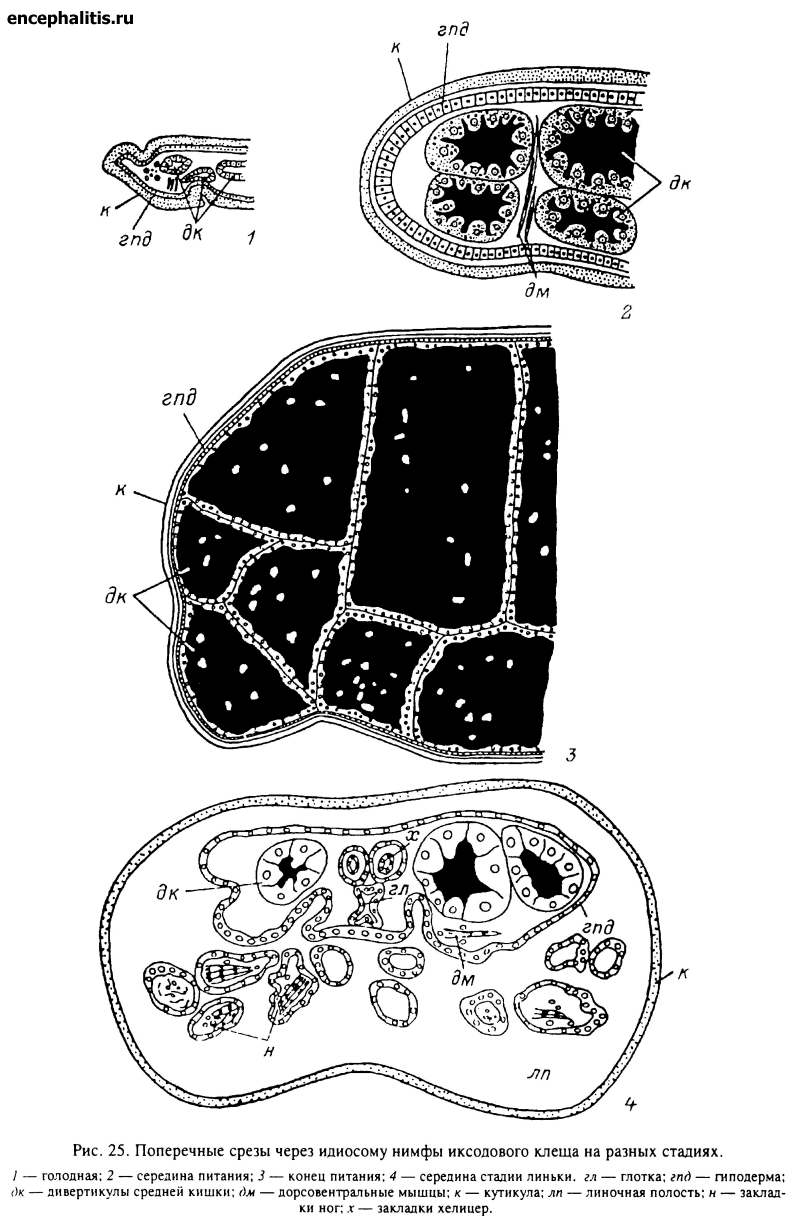
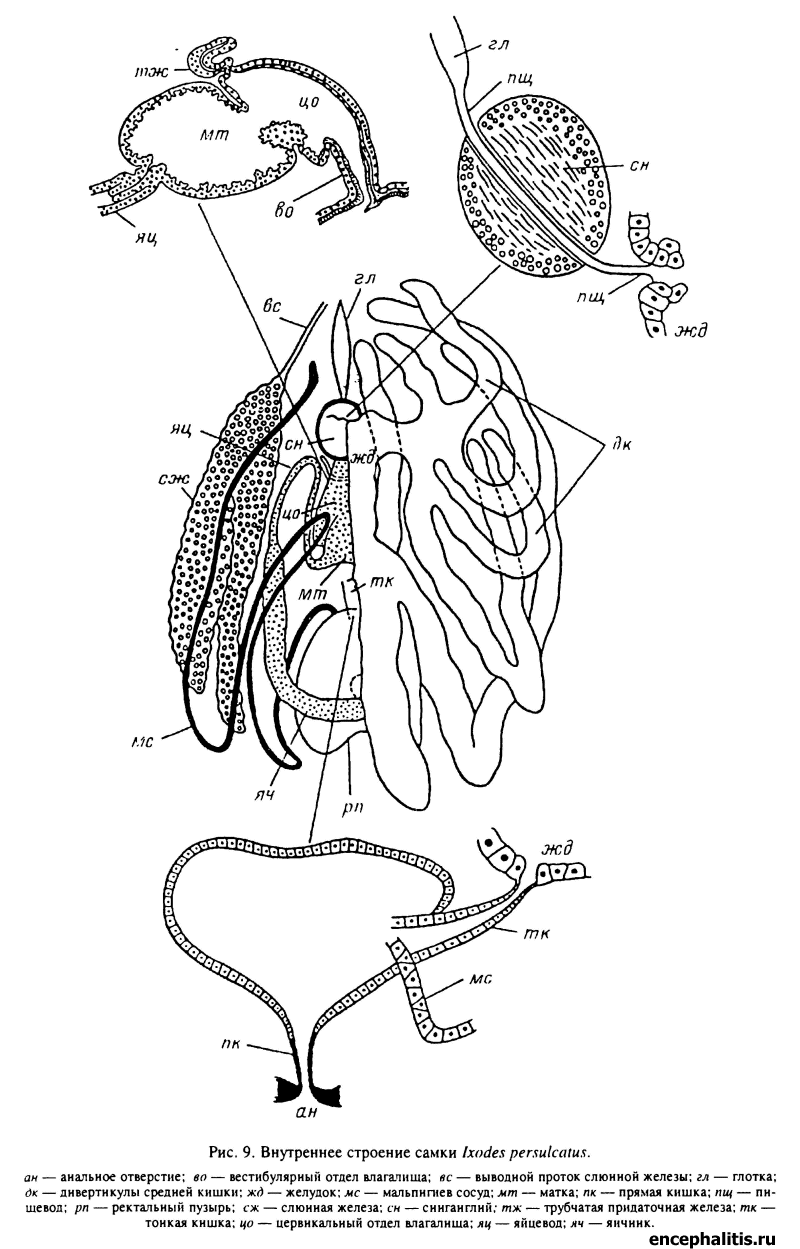
Средний отдел кишечника самки таежного клеща состоит из короткого трубковидного центрального отдела, или желудка, и трубчатых дивертикулов, расположенных сверху над остальными внутренними органами (рис. 9). В передневентральный конец желудка открывается пищевод, а с боков отходят длинные трубковидные боковые дивертикулы. При едином плане строения пищеварительной системы у всех иксодид в зависимости от видовой принадлежности, морфологической фазы, пола и стадии питания число, форма, размеры и взаимное расположение дивертикулов кишечника могут значительно варьировать. У самки таежного клеща (рис. 9) имеется 7 пар дивертикулов, из которых 1-я пара представлена короткими мешковидными выростами, а 2—5-я и 6—7-я пары сначала отходят короткими общими основаниями и затем разделяются на самостоятельные отростки. Отростки 1-й пары имеют форму коротких выростов. Наиболее длинные отростки 6—7-й пар, которые сначала идут в заднебоковом направлении, а затем подгибаются на вентральную сторону и доходят до гнатосомы; 2—5-я пары дивертикулов значительно короче, но тоже изгибаются на вентральную сторону. У голодных самок дивертикулы кишечника тонкие и свободно лежат между внутренними органами. По мере насыщения клеща во время питания желудок и особенно дивертикулы увеличиваются в размерах во много раз; они заполняют большую часть идиосомы, занимая все свободное пространство между внутренними органами.
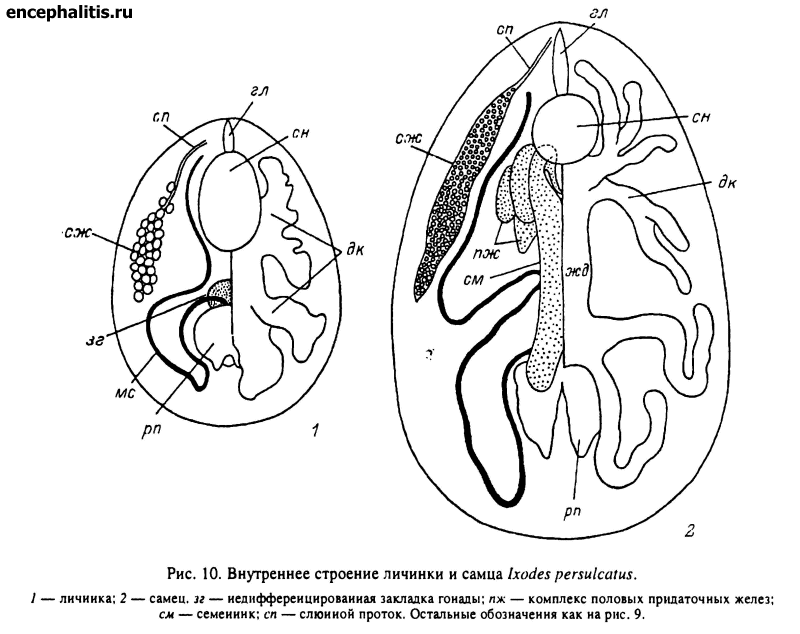
У личинки желудок более массивный, а отростки короткие, мешковидные рис. 10, 1). У нимфы форма средней кишки сходна с описанной для самки, но желудок массивнее, а боковые дивертикулы короче и толще. У самца общая форма кишечника сходна с таковой самки, но дивертикулы значительно короче и часть их может редуцироваться (рис. 10, 2).
От задневентрального конца желудка в задневентральном направлении отходит короткая трубка, называемая тонкой кишкой. Немного не доходя до вентральной стенки тела, она изгибается вверх и входит вместе с мальпигиевыми сосудами в передневентральную часть ректального пузыря (рис. 9). Диаметр тонкой кишки варьирует в зависимости от фазы развития клеща и степени заполнения: от тонкого клеточного тяжа у голодных личинок до сравнительно толстой трубки, заполненной кровью хозяина, у питающихся самок. Тонкая кишка является продолжением средней кишки и выстлана столбчатыми или кубическими эпителиальными клетками. Снаружи они окружены плексусом из продольных и кольцевых мышечных волокон.
Ректальный пузырь (рис. 9) представляет собой непарный мешок, форма и размеры которого в значительной степени зависят от степени заполнения гуанином и фекалиями. Стенки его построены из одного слоя плоских эпителиальных клеток, покоящихся на толстой базальной мембране и поверхностном мышечном слое. По своему строению клетки стенок ректального пузыря сходны с таковыми проксимальных отделов мальпигиевых сосудов и участвуют в процессах реабсорбции воды и солей.
Медиовентрально ректальный пузырь открывается в прямую кишку. Она опускается почти под прямым углом вниз и открывается анальным отверстием. На всем своем протяжении прямая кишка выстлана складчатой кутикулой, обеспечивающей возможность ее сильного растяжения при дефекации. От стенок прямой кишки к створкам анальной пластинки подходит пара мышц. При их сокращении анальная щель раскрывается, а нижняя часть прямой кишки немного выворачивается наружу. Сокращение кольцевой мускулатуры стенок тонкой кишки обеспечивает на этой стадии выталкивание фекалий наружу. По окончании акта дефекации кишка вворачивается внутрь, а створки анального отверстия замыкаются.
Несмотря на сложное морфологическое расчленение, все части средней кишки иксодид имеют сходное гистологическое строение. Строение кишечных клеток и особенности пищеварения были детально исследованы у клещей Н. asiaticum (Атлас..., 1979), I. persulcatus (Таежный клещ..., 1985), В. microplus (Agbede, Kemp, i985, 1987; Agyei et al., 1992), R. appendiculatus (Walker, Fletcher, 1986). Стенки кишки построены из однослойного эпителия, клетки которого покоятся на тонкой базальной мембране, снаружи оплетенной сетью мышечных волокон. Кишечный эпителий представлен 4 типами клеток: резервными, или камбиальными, секреторными, пищеварительными и желточными, соотношение между которыми значительно варьирует в зависимости от физиологического состояния клеща (рис. 12).
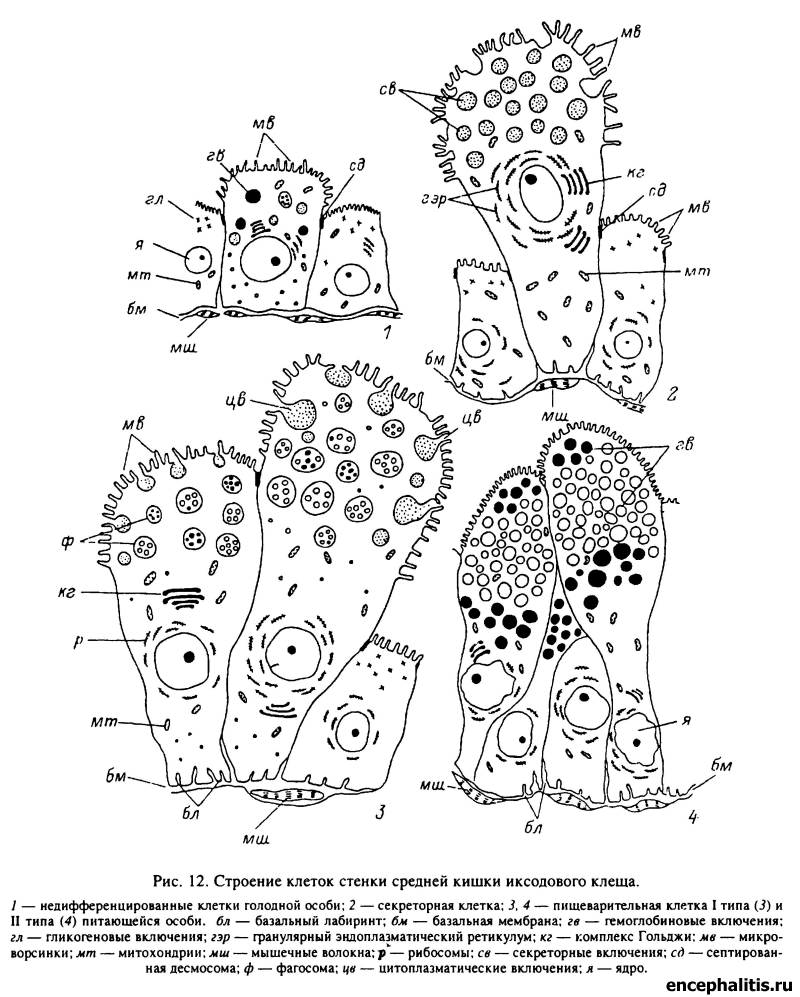
У голодающих особей эпителий средней кишки находится в состоянии «физиологической недоразвитости», что проявляется в отсутствии готовых к функционированию зрелых пищеварительных и секреторных клеток. Однако это ни в коей мере не исключает нормального протекания метаболических процессов в организме и связанного с ними расходования пищевых резервов клеток кишечника. По состоянию внутриклеточных пищевых включений, в частности, судят о физиологическом возрасте голодных клещей (см. главу V). Кишечный эпителий голодных клещей представлен в основном пищеварительными клетками предшествующей фазы развития и небольшим числом резервных клеток. Пищеварительные клетки нимфальной фазы в зависимости от физиологического возраста могут быть сильно гипертрофированными, с цитоплазмой, заполненной крупными пищевыми включениями у недавно перелинявших особей (табл. XX, 1, 2), или же сильно сжавшимися на разных стадиях дегенерации у длительно голодавших клещей (табл. XX, 7, 8).
Резервные клетки представляют собой камбиальные элементы, за счет деления и последующей дифференцировки которых происходит образование других типов кишечных клеток. Резервные клетки располагаются непосредственно у базальной мембраны. Они имеют цилиндрическую или кубическую форму и достигают у самок высоты 10—20 мкм (табл. XXI, 1). Для них характерны слабое развитие микроворсинок на апикальной поверхности и базальных впячиваний цитоплазматической мембраны, бедность цитоплазмы включениями и значительный объем ядра по отношению к цитоплазме.
Во время многодневного питания клещей происходят глубокие циклические изменения в клеточном составе кишечного эпителия. С самого начала прикрепления начинается пролиферация резервных клеток и дифференцировка их в секреторные и пищеварительные клетки. Первыми признаками их дифференцировки служат резкое увеличение гранулярной части ядрышка, развитие гранулярного эндоплазматического ретикулума и увеличение числа митохондрий (табл. XXII, 1).
В течение первых 1—2 сут питания значительная часть резервных клеток превращается в секреторные и в меньшей степени в пищеварительные. Секреторные клетки на этой стадии являются доминирующими элементами стенки кишечника. Для этих клеток характерна столбчатая или булавовидная форма при высоте 50— 70 мкм (рис. 12; табл. XXI, 2). Апикальная зона цитоплазмы сильно вакуолизирована. Секрет, заключенный в вакуолях, дает ШИК-положительную реакцию на углеводы и содержит основные белки. Цитоплазма этих клеток заполнена параллельными рядами цистерн гранулярного эндоплазматического ретикулума, большим количествам митохондрий и комплексов Гольджи. В апикальной зоне клеток появляется большое количество вакуолей с кислой фосфатазой. Освобождение клеток от секрета происходит по мерокриновому типу путем постепенного выброса секреторных гранул в полость кишки. Кроме того, часть секреторных клеток целиком отторгается в просвет кишечника и затем разрушается, сразу освобождая всю массу секрета (голокриновый тип секреции).
У иксодовых клещей преобладают процессы внутриклеточного пищеварения. У H. asiaticwn и I. persulcatus они осуществляются пищеварительными клетками I и II типов, которые являются основными компонентами стенки средней кишки. Апикальная поверхность этих клеток, обращенная в просвет кишечника, покрыта микроворсинками, а в базальной зоне лежит ядро и находятся впячивания цитоплазматической мембраны. Цитоплазма наряду с митохондриями содержит несколько типов эндоцитозных структур, пищевые включения, остаточные тела и другие элементы, связанные с осуществлением функций внутриклеточного пищеварения и метаболизма.
Пищеварительные клетки I типа преобладают на стадии медленного питания вплоть до осеменения самок. Они имеют столбчатую или булавовидную форму, достигают высоты 80—150 мкм, и их концы глубоко вдаются в полость кишечника (табл. XXI, 3). Для них характерна способность к поглощению компонентов пищи как путем пиноцитоза, так и фагоцитоза целых клеток крови и лимфы или их крупных фрагментов, которые затем подвергаются лизису внутри крупных фагосом. Часть этих клеток отторгается в просвет кишечника, где в них продолжаются процессы внутриклеточного пищеварения. Вследствие этой особенности достигается существенное увеличение поверхности кишечника, контактирующей с перевариваемой пищей.
Начиная со стадии быстрого насыщения пищеварительные клетки I типа постепенно исчезают. Процессы пищеварения сосредоточиваются во вновь образующихся из резервных пищеварительных клетках II типа. Для последних характерны очень крупные размеры, достигающие у самок в середине питания 100—150 мкм в высоту, а у готовящихся к откладке яиц — 400—500 мкм (табл. XXI, 4). Эти клетки поглощают питательные вещества путем пиноцитоза (табл. XXII, 4). Цитоплазма их заполнена включениями гемоглобина диаметром 10—15 мкм (табл. XXII, 3), а также более крупными жировыми включениями и остаточными телами, заключающими в себе гематиновые гранулы, пузырьки и мембраны. К началу яйцекладки клетки II типа поглощают всю кровь из полости кишечника. В результате просвет последнего сильно сужается, а апикальные концы гипертрофированных клеток соприкасаются между собой.
У заканчивающих яйцекладку самок большая часть цитоплазмы пищеварительных клеток бывает загружена гематином, а сами клетки находятся на разных стадиях дегенерации. Полость кишки на этой стадии опять несколько увеличивается и заполняется буро-черной кашеобразной массой из конкреций гематина, а также остатками дегенерировавших клеток. Общий объем дивертикулов заметно уменьшается за счет спадания их стенок, и они свободно лежат в полости тела.
После осеменения самок в стенках средней кишки образуются специализированные, так называемые желточные клетки, строение которых детально исследовано у самок D. variabilis (Tarnowski, Coons, 1989). Они имеют узкое основание, как бы вклинивающееся между соседними пищеварительными клетками. Апикальные части этих клеток, напротив, сильно расширены и частично прикрывают пищеварительные клетки. Для желточных клеток характерна высокая синтетическая активность, и в них формируется белок вителлогенин. Последний поступает в гемолимфу и из нее в развивающиеся ооциты, где он входит в состав желточных гранул. Максимальных размеров желточные клетки достигают у напитавшихся самок перед началом яйцекладки, после чего они постепенно спадаются и дегенерируют. У самцов и неполовозрелых фаз желточные клетки отсутствуют.
У личинок и нимф состав и строение клеток стенок средней кишки те же, что и у самок. Замена пищеварительных клеток на клетки следующей фазы происходит у них более растянуто и в значительной степени после линьки, по мере расходования заключенных в них пищевых резервов. Поэтому у недавно перелинявших особей кишечный эпителий образован главным образом пищеварительными клетками II типа предшествующей фазы развития.
Клеточный состав кишечника у самцов Amblyomminae тот же, что и у самок. Аналогичным образом протекают у них и процессы пищеварения. Однако в связи со значительно меньшей, чем у самок, массой поглощаемой пищи процессы ее переваривания протекают менее интенсивно и в кишечнике преобладают пищеварительные клетки I типа. У самцов таежного клеща и других видов Ixodes в связи с тем, что они в норме не питаются, цикл изменений в средней кишке связан с постепенным расходованием пищевых резервов нимфальной фазы и сходен с таковым голодающих самок.
У питающихся особей одновременно происходит как заполнение кишечника пищей, так и ее переваривание. Уже в пищевой полости и окружающей ее некротической зоне кровь, лимфа и другие компоненты пищи претерпевают первые деструктивные изменения. Они обусловлены как воздействием на ткани хозяина определенных компонентов слюны, так и гидролаз организма хозяина, которые освобождаются из нейтрофилов, мигрирующих к месту прикрепления клеща. Вследствие этих процессов в среднюю кишку поступает полностью или частично гемолизированная кровь, но на последней стадии питания в полость кишечника поступают и большие порции цельной крови с неповрежденными эритроцитами.
Следующий этап пищеварения связан с концентрацией жидкой пищи. Избыточная вода и соли выводятся из полости кишки в гемолимфу, а из нее с помощью специализированных клеток слюнных желез — в слюну и далее обратно в организм хозяина (см. главу VI). Поэтому в базальной зоне кишечных клеток особенно сильно развиты структуры лабиринта и межклеточные полости. Обезвоживание пищи происходит на протяжении всего периода питания, так что у личинки и нимфы таежного клеща концентрация крови хозяина в кишечнике увеличивается в 2—3 раза (Kitaoka, Fujisaki, 1976).
Дальнейшие этапы пищеварения протекают внутриклеточно в фаголизосомах пищеварительных клеток. Эти процессы происходят достаточно медленно и в случае диапаузы могут занимать несколько месяцев. На основании изучения нескольких видов иксодид (Akov, 1982) установлено, что протеазы клещей по своим свойствам близки к катепсину и имеют оптимум рН, близкий к 3. Столь кислая среда может достигаться только внутри фаголизосом, тогда как рН содержимого полости кишечника составляет около 6.5.
Следует отметить, что процессы пищеварения в отдельных клетках, даже расположенных поблизости друг от друга, а тем более в разных дивертикулах, не синхронизированы. В результате у питающихся или напитавшихся особей одновременно встречаются клетки на разных стадиях внутриклеточного пищеварения, сильно различающиеся по размерам, а также количеству и составу включений в цитоплазме.
У напитавшихся личинок и нимф значительная часть белков крови сохраняется внутри кишечных клеток в качестве пищевых резервов. Часть белков превращается внутри клеток в резервные липоидные включения. Именно описанные резервные белковые и жировые включения являются пищевым запасом длительно голодающих особей иксодид. У питающихся самок большая часть поглощенной пищи расходуется на развитие яиц. Процессы переваривания крови протекают у них быстрее, чем у неполовозрелых стадий. У напитавшихся самок D. variabilis за 12-суточный период от отпадения и до начала яйцекладки концентрация гемоглобина в организме снижается на 50 %, и этот процесс продолжается во время яйцекладки. Одновременно концентрация в кишечнике конечного продукта переваривания гемоглобина — гематина увеличивается к концу яйцекладки в 2 раза.
Средний отдел кишечника иксодовых клещей наряду со своей основной функцией — перевариванием пищи служит также в качестве депо запасных питательных веществ и местом их метаболических превращений. Соответственно он принимает на себя часть функций жирового тела и печени. Главным пищевым резервом у таежного клеща служат внутриклеточные включения гемоглобина, заполняющие значительную часть цитоплазмы пищеварительных клеток у недавно перелинявших особей. Наряду с усвоением белков крови продукты их переваривания могут подвергаться сложным метаболическим превращениям, и за счет этих процессов в клетках может накапливаться большое количество жировых включений. Последние, по-видимому, являются важным энергетическим резервом для голодающих особей.