Иксодовые клещи — паразиты и переносчики инфекций
Происхождение и эволюция
Проблемы происхождения и филогенетических связей иксодовых клещей из-за ограниченности палеонтологических находок базируются главным образом на косвенных данных из области сравнительной морфологии, биогеографии и паразито-хозяинных отношений.
Анализ списков хозяев разных видов клещей свидетельствует об отсутствии для многих из них строгих ограничений к переходу на новых прокормителей. Возможность неоднократной смены главных хозяев в процессе эволюции отдельных таксонов иксодид затрудняет выявление первичных хозяев.
Мы полагаем (Балашов, 1989а), что не меньшую, а может быть и большую роль, чем коэволюция с хозяевами, в развитии иксодид имели преобразования заселяемых ими ландшафтов, неоднократно проходившие за многомиллионный период существования этих клещей. Ареалы многих изученных в этом отношении видов могут совпадать с границами определенных биомов (хвойный и широколиственные леса, степи, разные типы саванн, субтропических и тропических лесов и т. п.), но не с видовыми ареалами их прокормителей. В процессе эволюции ландшафтов, а на более коротких этапах под влиянием сукцессии или антропогенных факторов клещи при сохранении пригодных для их существования условий внешней среды легко приспосабливаются к паразитированию на новых хозяевах, заменивших исчезнувших прокормителей. В частности, в антропогене за последние 5—10 тыс. лет произошла замена главных хозяев многих видов иксодид — диких копытных на вытеснивший их крупный и мелкий рогатый скот.
В связи с привязанностью клещей к определенным видам ландшафтов корректировку времени и места формирования отдельных таксонов иксодид может дать зоогеография. При использовании зоогеографических данных следует допустить ограниченную способность этих клещей к преодолению морских и океанических пространств, разделяющих континенты, и в меньшей степени гор и пустынь. Даже в случае межконтинентальных заносов иксодид с мигрирующими птицами, регулярно происходящих между Европой, Африкой и Азией, мигранты обычно неспособны к закреплению в необычных для них геобиоценозах. Для ареалов иксодид характерно постепенное расширение или сужение границ в ответ на соответствующие многолетние изменения биотических и абиотических факторов, а не взрывная экспансия. Исторические факторы в расселении иксодид отражают сложные процессы развития отдельных таксонов в условиях изоляции континентов и обмены фаун при соединениях континентальных платформ. Среди гипотез о происхождении иксодид широкое распространение получили взгляды Хугстрала (Hoogstraal, 1978; Hoogstraal, Kim, 1985), который вслед за Шульцем (Schulze, 1937) считает Ixodoidea первичными паразитами рептилий, возникшими в конце палеозоя или начале мезозоя около 200 млн лет назад в условиях тропического климата. Они обладали достаточно крупными размерами, имели в своем жизненном цикле 3 активные стадии (личинка, нимфа, имаго) и послужили общими предками современных Ixodidae и Argasidae. Иксодидный ствол первоначально разделился на 2 ветви. К одной из них в настоящее время наиболее близки современные примитивные виды Ixodes, а к другой — Аропотта, Amblyomma и Haemaphysalis. Несколько позднее в мелу от второй ветви обособилась ветвь Hyalomma, а роды Dermacentor, Rhipicephalus и Boophilus возникли не ранее замещения рептилий млекопитающими и птицами, т. е. в начале кайнозоя около 70 млн лет назад.
Согласно второй точке зрения, впервые высказанной Б. И. Померанцевым (1948а), первичными хозяевами примитивных Ixodidae были однопроходные и сумчатые млекопитающие, с которыми и в настоящее время связаны наиболее примитивные австралийские виды рода Ixodes. Дальнейшая эволюция этого единственного рода в подсем. Ixodinae была связана с переходом к паразитизму на плацентарных млекопитающих и птицах. Первичные этапы эволюции проходили в мезофильных стациях с богатой растительностью, на которой находились клещи, пассивно подстерегающие своих прокормителей. На более поздних этапах эволюции в процессе адаптивной радиации часть видов Ixodes вторично перешла к постоянному обитанию в гнездах и норах млекопитающих и птиц. Меньшая часть видов приобрела способность к паразитированию на имагинальной фазе на крупных копытных животных (I. ricinus, I. persulcatus, I. scapularis и др.), перейдя на них с мелких стационарно-логовищных млекопитающих.
Все остальные роды Ixodidae были объединены Померанцевым (1948а) в подсем. Amblyomminae. Предполагается, что первичными хозяевами этого, более прогрессивного и молодого таксона с самого начала были плацентарные млекопитающие и вторично часть из них (роды Аропотта и Amblyomma) перешла к паразитированию на рептилиях. Первоначально более примитивные группы этого подсемейства также были связаны с мезофильными лесными ландшафтами, в которых и в настоящее время обитает большая часть видов родов Haemaphysalis, Аропотта, Amblyomma. Виды родов Rhipicephalus, Hyalomma и Dermacentor приспособились к существованию в открытых пустынно-степных ландшафтах и питанию на обитающих в них крупных бродячих млекопитающих, в первую очередь копытных.
Точки зрения о первичности связей иксодовых клещей с млекопитающими и птицами придерживается и Н. А. Филиппова (1977). Она считает, что род Ixodes возник в мелу и первичными хозяевами этих клещей были уже существовавшие тогда птицы и примитивные млекопитающие. Первичные связи с птицами и особенно с морскими, по-видимому, имела эволюционная ветвь из подродов Ceratixodes, Scaphixodes, Pholeoixodes, Coxixodes. Вторая ветвь этого рода, представленная подродами Exopalpiger, Endopalpiger, Sternalixodes, специализировалась к паразитированию на сумчатых и ехиднах, и ее представители сохранились преимущественно в Австралии. Третья и наиболее прогрессивная ветвь возникла уже в палеогене и с самого начала эволюционировала в направлении пастбищного паразитизма (Ixodiopsis, Afrixodes, Partipalpiger, Trichotoixodes, Ixodes). В ходе своей эволюции часть видов третьей ветви с пастбищным паразитизмом вторично перешла к гнездово-норовому паразитизму, а некоторые из аборигенных австралийских видов приспособились к паразитированию на недавно завезенных в Австралию плацентарных млекопитающих.
Взгляды сторонников и противников первичности паразитарных связей иксодид с рептилиями, естественно, расходятся и в отношении времени происхождения этих кровососов, родственных отношений внутри семейства и обоснованности таксонов надвидового ранга. Общее признание получило только разделение клещей на 2 группы: Prostriata (Ixodinae) с единственным родом Ixodes и Metastriata (Amblyomminae), включающей все остальные роды.
Исключительная древность клещей, свободноживущие формы которых известны по палеонтологическим находкам с девона (Shear et al., 1984; Krivolutsky, Druk, 1986), и большая обособленность иксодид от других паразитиформных клещей, по мнению Балашова (1989а), не исключают возможности возникновения общей предковой группы аргасовых и иксодовых клещей уже в триасе. Во всяком случае 3 находки ископаемых иксодид из кайнозоя представлены формами, принадлежность которых к современным родам не вызывает сомнений. Из балтийских палеогеновых янтарей возраста в 35—40 млн лет описан Ixodes succineus (Weidner, 1964). Из олигоценовых отложений США возраста около 30 млн лет описан I. tertiarius, а в доминиканских янтарях из конца эоцена или олигоцена (возраст 30— 40 млн лет) — самец Amblyomma, похожий на A. testudineus, встречающийся и сейчас в этом регионе (Lane, Poinar, 1986). Этим же временем датируется и найденный в доминиканских янтарях аргасовый клещ Ornithodoros antiquus (Poinar, 1995). Вероятно, уже 30—40 млн лет назад существовали 2 рецентных рода иксодид из групп Prostriata и Metastriata и один из родов аргасовых клещей. Из более молодых находок в плиоценовых отложениях Европы следует отметить клеща из уха волосатого носорога, который по морфологическим особенностям был сходен с современным D. reticulatus и имел возраст в несколько миллионов лет (Schille, 1916). Следовательно, даже ограниченные палеонтологические данные подкрепляют гипотезу о достаточно раннем возникновении иксодид. Они также свидетельствуют о медленном темпе эволюции в двух морфологически примитивных родах Ixodes и Amblyomma, сохранивших свои важнейшие признаки на протяжении всего кайнозоя вплоть до настоящего времени.
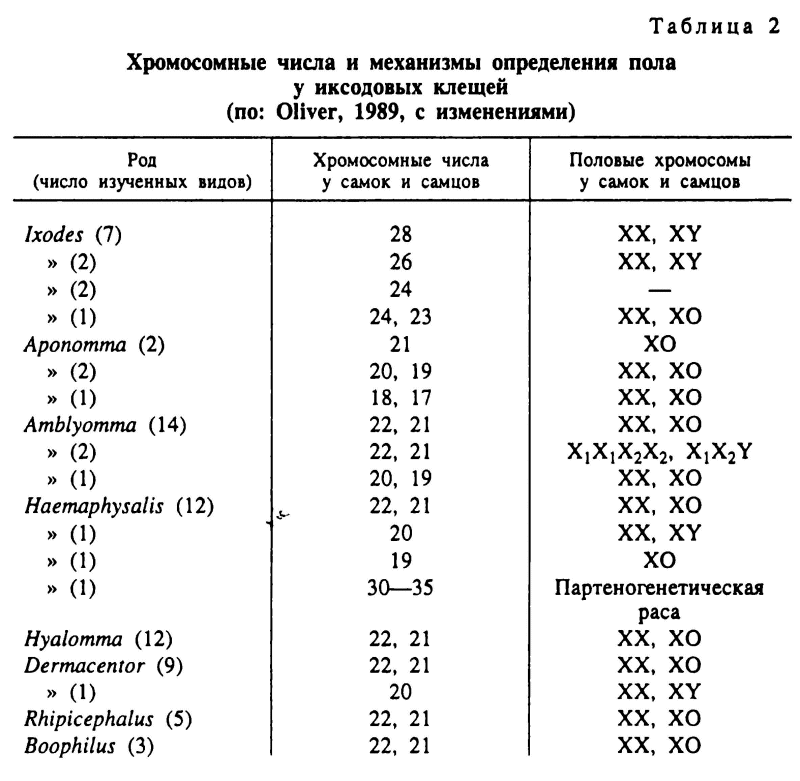
Важными критериями филогенетических отношений среди иксодид являются механизмы определения пола и кариотипы этих клещей (табл. 2) (Oliver, 1989). У всех Argasidae и среди иксодовых клещей у Prostriata с единственным родом Ixodes доминирует XY-механизм определения пола. Для подавляющей массы Metastriata определение пола осуществляется системой ХО. Лишь у немногих видов этой группы описаны также Х1 X2Y- и XY-системы половых хромосом. У Prostriata диплоидные наборы включают от 23 до 28 хромосом, и в этом отношении они ближе к аргасовым клещам с вариацией хромосомных чисел от 12 до 34. Особенно близки к роду Ixodes виды рода Argas с 26 хромосомами. В группе Metastriata у большинства видов самки имеют 22 и самцы 21 хромосому. Однако в родах Аропотта, Amblyomma и Haemaphysalis имеется несколько видов с иными хромосомными числами. Таким образом, цитогенетические данные подверждают обоснованность деления Ixodidae на две главные эволюционные ветви — Prostriata и Metastriata. Эти же данные указывают на большую близость Prostriata с Argasidae.
Иксодид первоначально разделили на Prostriata и Metastriata по внешним признакам. Не менее существенные различия наблюдаются между этими группами и в анатомических особенностях, включая выводные пути женской половой системы, типы секреторных клеток в слюнных железах, строение дермальных желез и присутствие в средней кишке Prostriata структуры, напоминающей перитрофическую мембрану насекомых. Во время линьки у видов Ixodes старая кутикула начинает разрываться сразу позади спинного щитка, а у Metastriata первый разрыв происходит между спинным щитком и гнатосомой. В процессе питания у всех изученных Metastriata ротовые органы клеща отделены от кожи хозяина застывшим секретом слюнных желез — цементным футляром. У многих Prostriata цементный футляр и аналогичные структуры, напротив, отсутствуют. Самцы Prostriata способны к спариванию и оплодотворению самок, не питаясь, а у многих видов они являются афагами. Сперматогенез происходит у них после нимфального питания на стадии подготовки к линьке (Балашов, 1956). У Metastriata для завершения сперматогенеза самцы нуждаются в питании, и только через несколько суток после его начала приобретают способность к спариванию. Исключение представляют несколько австралийских видов Аропотта, у которых голодные самцы имеют зрелые сперматиды и способны к осеменению самок (Oliver, 1989).
Существенный прогресс в установление степени генетического родства между современными таксонами иксодид внесло изучение последовательности оснований в митохондриальной рибосомной ДНК у 36 видов (Black, Piesman, 1994; Klompen et al., 1996). Эти данные свидетельствуют о существенных различиях между Prostriata и Metastriata, отражающих раннее расхождение этих эволюционных стволов. Среди Prostriata аборигенные австралийские виды, относимые к подродам Coxixodes, Ceratixodes, Stemalixodes и Exopalpiger, существенно отличаются по строению ДНК от всех остальных подродов рода Ixodes. Metastriata представляют монофилетический ствол. Внутри этого обширного таксона считающиеся более примитивными роды Аропотта и Amblyomma являются парафилетическими группами по отношению к Haemaphysalis и остальным родам (Dermacentor, Hyalomma, Rhipicephalus, Boophilus и др.), образующим более молодую ветвь развития иксодид.
Опираясь на изложенные выше данные, мы полагаем, что уже на раннем этапе своего эволюционного развития ствол проиксодид разделился на две главные ветви — предков аргасовых и иксодовых клещей. Для первых с самого начала был характерен гнездово-норовый тип паразитизма. Иксодиды эволюционировали в направлении усиления не только трофических, но и других паразитарных связей со своими прокормителями. Не исключено, что в процессе эволюции иксодоидных клещей возникали и другие филогенетические ветви, не оставившие следов, из которых в настоящее время обнаружен один вид Nutalliella namaqua, единственный в сем. Nutalliellidae, несущий в своей организации черты как аргасовых, так и иксодовых клещей (Roshdy et al., 1983).
Допуская древность происхождения иксодид, нам тем не менее кажется недостаточно обоснованным рассматривать рецентных представителей родов Аропотта и Amblyomma прямыми потомками паразитов пермских рептилий. По своим морфофизиологическим особенностям они достаточно продвинуты и обладают всеми характерными чертами временных эктопаразитов с длительным питанием. Рептилии являются единственными хозяевами только для видов рода Аропотта. В роде Amblyomma основную массу видов составляют паразиты нескольких отрядов млекопитающих.
Вообще гипотеза о непосредственном переходе иксодид с вымирающих крупных рептилий на млекопитающих недостаточно убедительна. Известно, что во время массового вымирания рептилий в меловой период крупные млекопитающие отсутствовали или были относительно редки. Они стали доминирующими жизненными формами только с конца палеоцена—начала эоцена. В то же время на протяжении значительной части мезозоя вместе с рептилиями сосуществовали мелкие однопроходные и сумчатые млекопитающие, имевшие постоянные норы, убежища или логовища, а также гнездовые колонии юрских птиц. Поэтому вполне допустимо, что некоторые клещи — паразиты рептилий еще при жизни своих первичных хозяев приспособились к паразитированию на новых для них классах прокормителей и стали предками современных видов рода Ixodes. Возможно, что сходным путем, но несколько позднее и уже в связи с плацентарными млекопитающими происходила эволюция рода Haemaphysalis, из 147 видов которого только один является строго специфичным паразитом сумчатых и еще для нескольких видов они служат дополнительными хозяевами. Не исключено также, что паразитирование неполовозрелых фаз развития 12 видов этого рода, и в их числе наиболее примитивного подрода Alloceraea, на рептилиях отражает их прошлые глубокие связи с этой группой прокормителей. В пользу докайнозойского возникновения родов Ixodes и Haemaphysalis также указывает их всесветное распространение.
У всех исследователей не вызывает сомнения сравнительная молодость родов Rhipicephalus, Dermacentor, Boophilus, Margaropus, связанных в своей эволюции с копытными животными, хотя взгляды на место и время их происхождения и родственные связи между ними неоднозначны. Более спорен вопрос о происхождении и эволюции рода Hyalomma, из 30 видов которого только 1 — Н. aegiptium на всех фазах развития паразитирует на черепахах, все остальные — паразиты млекопитающих. Взрослые клещи питаются преимущественно на диких и домашних копытных, а неполовозрелые фазы — кроме копытных на мелких и средних млекопитающих и редко (Н. marginatum) на птицах. Хугстрал (Hoogstraal, 1978) полагал, что род Hyalomma возник в меловой период, его первоначальными хозяевами были крупные рептилии группы Rhynchosauria. Позднее эти клещи перешли к паразитированию на крупных млекопитающих, и дальнейшая их эволюция протекала в связи с копытными в аридных ландшафтах. Мы думаем, что исходная предпосылка этой гипотезы, а именно первичность и древность связей Н. aegyptium с черепахами, недостаточно убедительна. В условиях степей и пустынь многие виды животных переходят к норовому образу жизни. Вероятно, подобный переход был возможен и у предков Н. aegyptium, для которых черепахи, многочисленные в пустынных ландшафтах, стали главными прокормителями. При этом личинки и нимфы сохранили способность и часто питаются на многих аборигенных видах млекопитающих и птиц. Более аргументированной поэтому представляется гипотеза Померанцева (19486), связывающая происхождение Hyalomma со временем формирования азиатских пустынь. В пользу этих предположений говорит полное отсутствие видов этого рода в Австралии и Америке, а также значительное сходство ДНК Hyalomma с таковой видов Dermacentor, Boophilus и Rhipicephalus.
Большая древность иксодид, отсутствие палеонтологических данных, а главное, очевидная легкость смены хозяев в процессе эволюции затрудняют, по нашему мнению, объективную оценку родственных отношений между рецентными таксонами. Эволюционные преобразования можно с достаточной обоснованностью оценить только на уровне родов, большая часть которых признается всеми иксодологами и, вероятно, отражает определенные эволюционные стволы большего или меньшего масштаба. В то же время предлагаемые надродовые и внутриродовые таксоны, за исключением разделения иксодид на 2 главных ствола — Prostriata (=Ixodinae) с единственным родом Ixodes и Metastriata (=Amblyomminae), включающий все остальные роды, представляются дискуссионными.