Иксодовые клещи — паразиты и переносчики инфекций
Описания жизненных циклов клещей рода Boophilus
Всем 4 видам этого рода свойствен однохозяинный тип развития. Наиболее детально он изучен у В. microplus, первоначальной родиной которого была Юго-Восточная Азия и откуда он в исторический период распространился по всей тропической и частично влажной субтропической зонам. У В. microplus в условиях постоянной высокой температуры и влажности, характерных для тропического климата Бразилии, Кубы или Северной Австралии, от отпадения напитавшихся самок до появления активных голодных личинок проходит всего около 4 нед. Если к этому добавить 3—4 нед питания и линек на теле хозяина, то время развития 1 генерации составляет около 8 нед. Фактические сроки жизни одного поколения увеличиваются в зависимости от сроков встречи с прокормителем голодных личинок (предельный срок жизни до 107 сут) еще на 1—4 нед. В среднем за год успевает развиваться 5—6 последовательных генераций клещей. Однако в случае с непрерывным циклом развития само понятие генерации представляется крайне неопределенным. Одновременно на теле хозяина находятся клещи разных фаз развития, на разных сроках питания. В связи с возможностью жизни голодных личинок до 1—2 мес питающиеся на одном хозяине и в одно время клещи могут принадлежать к родительскому и дочернему поколениям. Отпадение напитавшихся самок круглый год при отсутствии синхронизирующих факторов для сроков яйцекладки, эмбриогенеза и активности личинок в еще большей степени стирает всякие границы между поколениями. Реально в условиях тропического климата природные популяции В. microplus представлены совокупностями особей разных календарных возрастов без каких-либо генеративных разрывов.
В субтропическом климате прохладная зима прерывает или сильно замедляет развитие непаразитических стадий, а весной и осенью для их завершения требуется 10—12 нед. В Аргентине в подобных условиях в течение 1 года успевают развиться 3—4 поколения. Границы между ними обнаруживаются по нескольким последовательным пикам отпадения напитавшихся самок. Наиболее выражен первый весенний пик, который обусловлен одновременным нападением активизировавшихся с весенним потеплением личинок. Последующие пики становятся более пологими из-за увеличения различий в сроках развития непаразитических стадий в отличающихся по микроклимату местообитаниях. Десинхронизация развития связана также с разной продолжительностью жизни голодных личинок до встречи с прокормителем. В конце осени—начале зимы развитие напитавшихся самок и их кладок замедляется или прекращается, голодные личинки становятся неактивными и происходит частичное или полное освобождение скота от клещей (Nunez et al., 1985; Sutherst et al., 1988a).
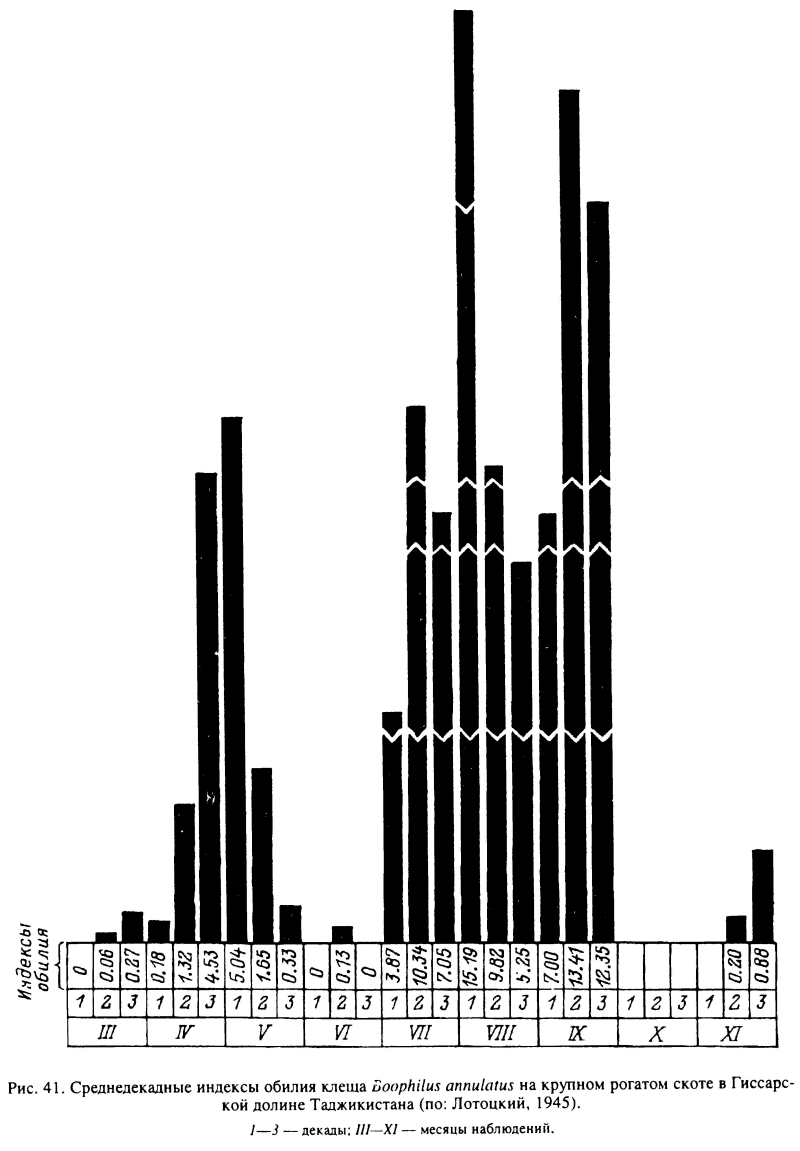
Сходный тип развития свойствен имеющему средиземноморское происхождение В. annulatus. На юге Казахстана и в Средней Азии зимние температуры ниже 5—10 °С, полностью исключающие возможность развития этих клещей, сохраняются в течение 3—4 мес. В этот период на пастбищах перезимовывают голодные личинки, напитавшиеся самки и яйца. Состояние диапаузы, по-видимому, не возникает, так как при достаточно теплой погоде в отдельные зимние дни наблюдалось нападение на скот активизировавшихся личинок в Южном Таджикистане. Как правило, зимой скот свободен от клещей. Массовое нападение личинок начинается с марта, и в течение теплого сезона на скоте наблюдаются весенняя, летняя и осенняя волны заклещевления (рис. 41). Последние, по-видимому, отражают смену 3 частично перекрывающихся поколений В. annulatus, успевающих развиться за 1 год (Потоцкий, 1945; Галузо, 1950). Для В. decoloratus, являющегося эндемичным паразитом диких полорогих и крупного рогатого скота в Южной Африке, в условиях местного субтропического климата за 1 год успевает развиться от 3 до 4 генераций. Им соответствуют 4 пика численности этих клещей на скоте в июле, сентябре—октябре, декабре—январе и марте—апреле (Rechav, Kostrezewski, 1991).