Иксодовые клещи — паразиты и переносчики инфекций
Описания жизненных циклов клещей рода Hyalomma
Для видов этого рода описаны в основном 1-годичные циклы, которые сохраняются даже у видов, обитающих в условиях пустынь Средней Азии и Казахстана с достаточно суровой и продолжительной зимой. В лабораторных культурах при температуре 25 °С средние сроки развития у 5 видов этого рода варьировали от 98 до 196 сут (Ouhelli, 1994).
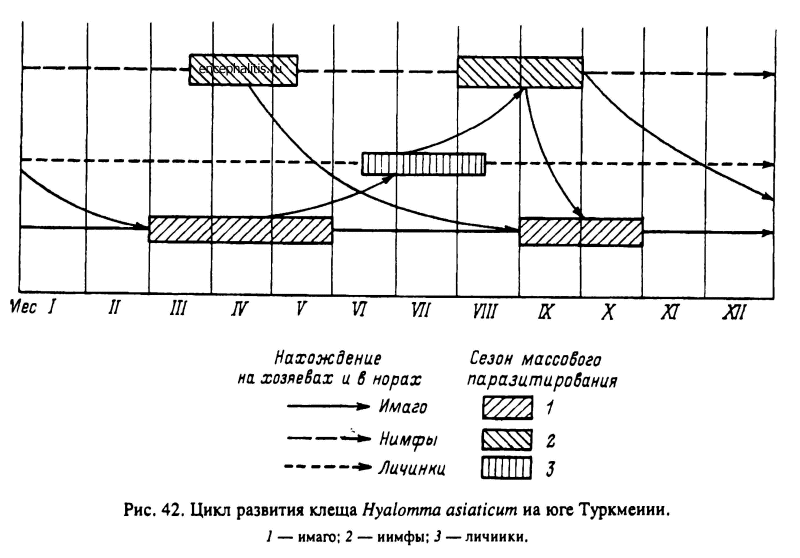
Н. asiaticum широко распространен в пустынных и полупустынных ландшафтах от Сирии на западе до Монголии и Центрального Китая на востоке, от Казахстана на севере до побережья Персидского залива на юге. Это треххозяинный вид, все фазы развития которого в пустынях Туркмении встречаются круглогодично. Массовое паразитирование имаго на скоте и верблюдах наблюдается в апреле—мае и сентябре—октябре. Личинки паразитируют на больших песчанках и других мелких и средних млекопитающих с марта по сентябрь с максимальной численностью в июне—июле, нимфы — на тех же хозяевах с февраля по ноябрь с максимумом в августе—сентябре. Напитавшиеся весной самки в условиях высоких температур без задержки приступают к яйцекладке, а вылупившиеся из яиц личинки дают пик численности на хозяевах в июне—июле. С перелинявшими на них нимфами связан позднелетний пик численности на песчанках. На зимовку уходят голодные и напитавшиеся имаго и нимфы и голодные личинки от позднелетних кладок. Значительная индивидуальная долговечность голодных особей, достигающая в условиях микроклимата нор большой песчанки нескольких месяцев для личинок и нимф и более 1 года для имаго, приводит к частичному смешиванию потомства от самок весеннего и осеннего питания и даже клещей двух последовательных генераций. Таким образом, при преимущественно 1-годичном цикле часть популяции, представленная вылупившимися осенью личинками, уже принадлежит второй генерации клещей (рис. 42). Развитие ее завершается уже после зимовки в следующем сезоне. У нимф же и особенно имаго, не сумевших в теплый сезон найти хозяина и ушедших на зимовку, развитие затягивается до 1.5—2 лет (Балашов, 1972; Бердыев, 1980).
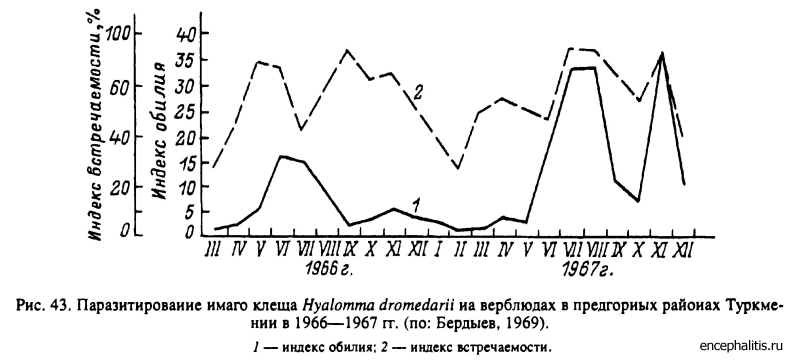
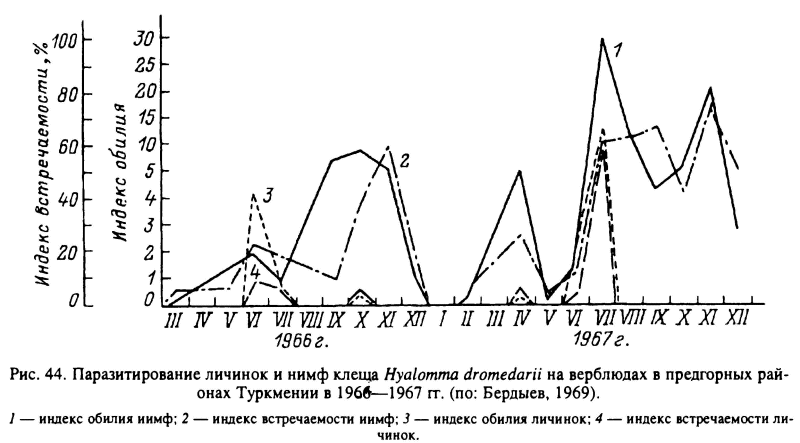
Н. dromedarii широко распространен в пустынных районах Африки к северу от экватора, в странах Ближнего и Среднего Востока, Средней Азии. Главными прокормителями для всех фаз служат верблюды и крупный рогатый скот. В южных частях ареала развитие идет преимущественно по треххозяинному типу, а на севере ареала в Туркмении в летние месяцы — по двух- и треххозяинному типам, а в зимние — по однохозяинному. В Туркмении взрослые клещи встречаются на верблюдах круглый год, но в максимальном количестве — в июне—августе и в ноябре (рис. 43). Нимфы также паразитируют на верблюдах круглогодично с максимумами в августе и ноябре (рис. 44), причем напитавшиеся в конце осени особи остаются зимовать на теле хозяина и отпадают только весной. Пики численности личинок на хозяевах наблюдаются в июне—июле и осенью (рис. 44). Напитавшиеся осенью самки впадают в зимнюю диапаузу, зимуют в почве и откладывают яйца весной. При треххозяинном типе развития для завершения 1 поколения в теплое время года требуется 103—127 сут, а при двух- и однохозяинном развитии эти сроки еще короче. Соответственно, с учетом зимней остановки, в течение 1 года успевает развиться до 1.5 поколения и более, а границы между последовательными поколениями стерты (Бердыев, 1969, 1980).
Ареал Н. anatolicwn охватывает пустынные и степные ландшафты Средиземноморья и Северной Африки почти до экватора, Аравийский полуостров, Иран, Пакистан, Индию и Среднюю Азию. Клещи при питании всех фаз на крупном рогатом скоте имеют преимущественно двуххозяинный цикл, а при питании личинок на мелких млекопитающих — треххозяинный. На юге Туркмении имаго паразитируют на скоте круглогодично. Численность их быстро возрастает с середины апреля и достигает максимума в июне—августе, затем постепенно снижается, так что с ноября по март на животных встречаются единичные особи (преимущественно самцы). Личинки паразитируют с мая до конца сентября с максимумом в августе, нимфы — с мая по октябрь с максимумом в июле. Зимуют голодные и напитавшиеся имаго и нимфы и, возможно, в небольшом количестве личинки. Перезимовавшие и активизировавшиеся весной имаго сразу после питания приступают к яйцекладке одновременно с перезимовавшими самками осеннего насыщения. Вылупившиеся из весенних кладок личинки успевают к концу осени развиться до стадии голодных или напитавшихся нимф, которые уходят на зимовку. Перезимовавшие сытые и голодные нимфы уже в конце мая—июне линяют в имаго, которые успевают напитаться или уходят на зимовку голодными. В результате продолжительность развития 1 поколения занимает от 1 до 1.5—2 лет (Бердыев, 1980).
Сходный 1-годичный цикл развития описан у этого вида и в Израиле (Rubina et al., 1982). В условиях эксперимента показано (Белозеров, 1981), что под влиянием длины дня может возникать состояние диапаузы. Она проявляется в задержке развития напитавшихся нимф, оогенеза самок и развития яиц, а также в снижении агрессивности и увеличении срока выживания непитавшихся особей. В условиях Туркмении диапауза возникает с начала сентября при длине дня в 13 ч и менее, а с середины сентября она наблюдается у 100 % напитавшихся в природе нимф. В Египте формирование диапаузы этого вида происходит позднее, когда среднесуточные температуры опускаются ниже 25 °С. У напитавшихся самок диапауза возникает при длине дня в 14 ч и меньше, и соответственно в условиях Средней Азии задержка откладки яиц наблюдается у особей, напитавшихся в конце августа или начале сентября. В Египте же уход самок на зимовку зависит главным образом от температуры, и диапауза может возникнуть не ранее конца сентября—начала октября. Существование стадии морфогенетической диапаузы у нимф и самок H. anatolicum обеспечивает сезонную упорядоченность цикла развития этого вида за счет совпадения сроков линьки нимф или яйцекладки самок независимо от осеннего или весеннего периодов их питания.
Н. detritum имеет сходное с предыдущим видом распространение, но заселяет более мезофильные местообитания и тяготеет к орошаемой зоне, долинам рек.
Цикл развития двуххозяинный, и все стадии питаются преимущественно на крупном рогатом скоте. При облигатном моновольтинном цикле развития за год развивается только 1 поколение клещей, даже если сумма эффективных температур достаточна для частичного развития второй генерации. Диапауза в этих случаях не только обеспечивает перезимовку клещей, но и синхронизирует их жизнедеятельность с определенными фенологическими сезонами. В условиях Южного Казахстана и в Средней Азии имаго паразитируют на скоте в июне—августе. Напитавшиеся самки через 1—2 нед приступают к яйцекладке, и из яиц без задержки вылупляются личинки. Последние остаются скоплениями в местах кладок в неактивном состоянии до конца сентября—октября. С начала октября начинается нападение личинок на скот, и напитавшиеся нимфы отпадают в октябре—ноябре. Часть нимф остается на теле хозяев и отпадает с началом весны в марте. Независимо от сроков отпадения линька нимф происходит в конце весны, и свеже-перелинявшие имаго начинают без задержки нападать на скот (Галузо, 1947; Бер-дыев, 1980).
Н. scupense сходен в своем распространении с Н. detritum, но заходит дальше на север, встречаясь на юге Украины и России. Развитие идет только по однохозяинному циклу. Личинки нападают на скот не ранее конца сентября. Напитавшиеся личинки остаются на теле хозяина и линяют в нимф, без задержки продолжающих питание. Имаго, перелинявшие из нимф в декабре—феврале, остаются прикрепившимися к хозяину, но не питаются, по-видимому находясь в состоянии диапаузы. Первые напитавшиеся самки в условиях Южного Казахстана появляются с конца марта, и отпадение их продолжается в апреле—начале мая. Они без задержки откладывают яйца, из которых в конце весны—начале лета вылупляются личинки. Последние весь летний период остаются в почве и растительных остатках в неактивном состоянии. Активные личинки поднимаются на растительность и нападают на скот только с конца сентября (Галузо, 1947).
Н. marginatum обладает исключительно обширным ареалом, включающим Средиземноморье, Среднюю Азию, большую часть Африки и Индостанский полуостров. Клещи встречаются в различного типа степях и саваннах, включая аридные горные редколесья. Цикл развития двуххозяинный. Неполовозрелые фазы паразитируют на питающихся на земле птицах, зайцах, ежах и других млекопитающих, а взрослые клещи — на крупном рогатом скоте, лошадях и других домашних и диких животных с крупными и средними размерами тела. В Средней Азии, Закавказье и на юге России цикл развития 1-годичный моновольтинный. Зимующими стадиями являются голодные имаго и напитавшиеся нимфы. Активация перезимовавших или перелинявших весной имаго и нападение их на млекопитающих происходят весной и в первой половине лета. Напитавшиеся самки без задержки откладывают яйца, из которых через 20—40 сут вылупляются и сразу же активизируются личинки. Сезон паразитирования их ограничен второй половиной лета и осенью (Галузо, 1947; Березин, 1971; Бердыев, 1980).
Н. aegyptium, паразитирующий на всех фазах преимущественно на наземных черепахах в Средиземноморье, Закавказье, Иране и Средней Азии, обладает треххозяинным 1-годичным циклом развития. Весной на активизировавшихся после зимней спячки черепахах последовательно сменяются питающиеся личинки, нимфы и в начале лета — имаго.