Иксодовые клещи — паразиты и переносчики инфекций
Описания жизненных циклов клещей рода Haemaphysalis
Несмотря на большое видовое разнообразие, жизненные циклы изучены только для немногих видов. Всем им свойствен треххозяинный тип развития продолжительностью от 1 до 4 лет.
Наиболее детально изучено развитие Н. longicornis, обитающего в Юго-Восточной Азии в странах с субтропическим и умеренным климатом, включая и юг Приморского края в России. В связи с обширным ареалом эти клещи встречаются в разных типах лесов и на травянистых пастбищах при условии их достаточной увлажненности. Взрослые паразитируют на многих видах травоядных и хищников, особенно на оленях и на скоте. Неполовозрелые питаются на мелких млекопитающих, козах и на птицах. В зависимости от климата цикл развития может быть 1-годичным на юге и 2—3-годичным на севере. В Австралии (Sutherst, Bourne, 1991) в более прохладных горных лесах зимуют диапаузирующие голодные взрослые и неполовозрелые клещи; жизненный цикл клеща хорошо синхронизирован со сменой сезона года и завершается за 1 год. В более жарких прибрежных местообитаниях у части популяции осенне-зимняя диапауза нарушается, и развитие может быть более коротким. В Японии и Южной Корее с более холодной зимой цикл развития 2-годичный с зимовкой голодных имаго и нимф (Ito et al., 1983). В Приморском крае у северной границы ареала (Беликова, 1969) цикл развития становится 2- и 3-годичным с зимовкой голодных личинок, нимф и взрослых.
У партеногенетической расы Н. longicornis голодные личинки и нимфы при 18 °С были активны - в условиях длинного дня (14—24 ч света). При освещенности менее 13 ч у личинок и 15 ч у нимф у клещей возникала и поддерживалась свыше 4—5 мес поведенческая диапауза в форме отсутствия агрессивности. Перенос клещей в условия длинного дня вызывал в течение 1—3 нед активацию, и клещи начинали нападать на прокормителей. У напитавшихся личинок фотопериодические условия также регулируют продолжительность метаморфоза, так что в коротком дне развитие шло быстрее, чем в длинном (Белозеров, Лузев, 1974). Фотопериодическая индукция диапаузы голодных особей обнаружена в природе в Австралии, где в условиях укорачивающегося дня голодные нимфы и имаго становились неактивными (Sutherst, Bourne, 1991). В Японии нахождение голодных нимф и имаго этого вида в условиях короткого дня также вызывало развитие поведенческой диапаузы, а развивающиеся яйца, голодные и напитавшиеся личинки не реагировали на длину дня (Fujimoto, 1995a).
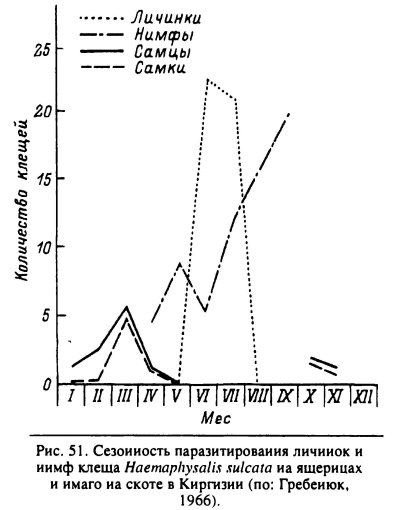
Н. sulcata распространен в степных и полупустынных ландшафтах Средиземноморья и Средней Азии. Взрослые паразитируют главным образом на козах и овцах. Неполовозрелые питаются на многих видах рептилий, реже на птицах и мелких млекопитающих. В Израиле (Feldman-Muhsam, Borut, 1983b) в условиях мягкой зимы взрослые паразитируют без перерыва с ноября по апрель. Самки, закончившие питание в начале зимы, приступают к яйцекладке в марте—апреле вместе с особями от более поздних сроков отпадения. Личинки появляются в мае и встречаются на рептилиях до августа, часто вместе с нимфами. Напитавшиеся личинки вскоре линяют в нимф, и последние питаются с конца июня до сентября. Напитавшиеся нимфы успевают перелинять в имаго осенью и сразу же начинают нападать на хозяев. В результате развитие 1 генерации укладывается в 1 год. В Киргизии (Гребенюк, 1966) с достаточно суровой зимой цикл развития занимает 1.5—2 года. Взрослые паразитируют на скоте преимущественно осенью и ранней весной с зимним перерывом. Неполовозрелые встречаются на рептилиях весь летний период (рис. 51). Напитавшиеся личинки успевают в этом же теплом сезоне перелинять в нимф, но только часть из последних успевает напитаться до наступления осенних холодов. Значительная часть голодных и все напитавшиеся нимфы зимуют и завершают развитие следующим летом. Перелинявшие из них имаго зимуют напитавшимися или голодными и откладывают яйца только после зимовки.
Н. punctata также принадлежит к числу широко распространенных видов и ареал его сходен с таковым Н. sulcata, но заходит дальше на север, включая некоторые острова Северного и Балтийского морей. Вид обладает высокой экологической пластичностью и встречается в разных типах равнинных и горных ландшафтов со степной, кустарниковой и разреженной лесной растительностью. Список хозяев насчитывает несколько десятков видов млекопитающих, а для неполовозрелых фаз — также птиц и рептилий. Активность имаго в зависимости от географической широты местности имеет 2 пика, различающихся по продолжительности и обилию клещей, — весенний и осенний. Неполовозрелые фазы активны с начала весны и до конца осени (Гребенюк, 1966; Nosek, 1973; Успенская, 1987). В Молдавии зимуют все фазы развития и продолжительность жизненного цикла сильно варьирует. В оптимальных условиях развитие 1 генерации завершается за 1.5 года. Потомство напитавшихся весной самок успевает в этом же сезоне достигнуть стадий голодных или напитавшихся нимф, которые после зимовки уже к середине следующего лета достигают имагинальной фазы. Преобладает 2-годичный цикл, когда от весенних кладок яиц на зимовку уходят напитавшиеся или голодные личинки, которые только с середины следующего лета линяют в нимф. Последние зимуют голодными или напитавшимися и на следующее лето линяют в самцов и самок, способных к осеннему питанию. У части популяции клещей, не сумевших напитаться ранее второй половины осени, развитие 1 поколения может затягиваться до 3 и даже 4 лет. Во всех случаях при прочих равных условиях отмечено сокращение сроков эмбрионального развития и линек личинок и нимф в весенне-летний период и удлинение их в осенние месяцы.
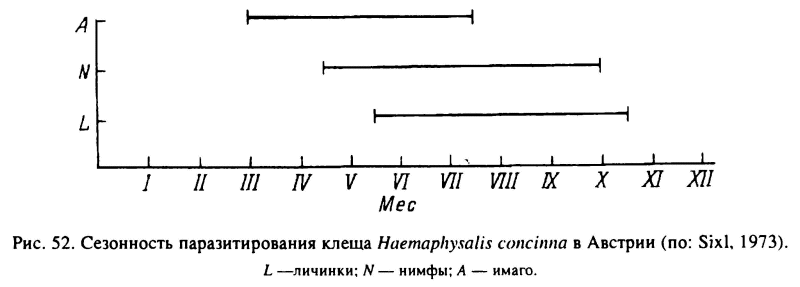
Н. concinna в своем распространении связан с поясом широколиственных лесов Палеарктики. Кроме разных типов мезофильных лесов клещи встречаются на влажных лугах, в зарослях кустарника и камыша. Хозяевами служат многие виды диких млекопитающих, скот и птицы. Повсеместно на протяжении теплого времени года одновременно активны на растительности и паразитируют все фазы развития (рис. 52). Зимуют также все фазы, кроме напитавшихся самок и яиц. Развитие 1 генерации продолжается 3 года и более (Беликова, 1969; Nosek, 1973; Lebedeva, Korenberg, 1981). Синхронизация жизненного цикла в условиях больших климатических различий в границах ареала достигается поведенческой диапаузой голодных взрослых и неполовозрелых особей, а также морфогенетической диапаузой у напитавшихся личинок и нимф. У личинок короткий день обеспечивает длительное, свыше 3 мес сохранение их скоплений в местах вылупления. В условиях длинного дня личинки активизируются и расползаются от места кладки уже через 10 сут. У нимф и взрослых длина дня непосредственно контролирует их агрессивность. В эксперименте в условиях длинного дня к мышам присасывалось 41—58 % выпущенных на них нимф, а в коротком дне только 5—7 % (Белозеров, 1974).
Для Н. bispinosa, обитающего в тропическом климате Индийского субконтинента от Шри-Ланки до Гималаев, характерен 1-годичный цикл развития. Питание взрослых, яйцекладка, развитие личинок и нимф завершаются в сезон летних муссонных дождей, а сухой сезон с декабря до мая переживают неактивные голодные имаго (Bhat, 1971).
Высокой экологической пластичностью отличается Н. leporispalustris. Этот вид распространен от Аляски до Аргентины, где приурочен к местообитаниям своих главных прокормителей — зайцеобразных. Кроме кроликов и зайцев, на которых питаются все фазы развития, неполовозрелые в большом количестве паразитируют на многих видах птиц. В южных прибрежных районах США с мягким климатом цикл развития 1-годичный. Личинки в наибольшем количестве встречаются на хозяевах с июля по декабрь, нимфы — в августе—сентябре, а взрослые — весь год с максимумом в мае (Sonenshine, Stout, 1970). В районах с холодной зимой, как например в Миннесоте (Green et al., 1943), с декабря по март пораженность клещами их главного хозяина — зайца-беляка минимальна, так как зимующие голодные особи всех фаз развития находятся в неактивном состоянии. Количество питающихся клещей начинает увеличиваться только с апреля, и большое количество питающихся на всех фазах сохраняется с мая по июль. Быстрое снижение питающихся особей происходит в октябре—ноябре. В мае в наибольшем количестве паразитируют взрослые, а в августе — личинки. Как можно судить по сезонности активности неполовозрелых и взрослых особей, для развития 1 генерации требуется не менее 2 лет.