Иксодовые клещи — паразиты и переносчики инфекций
Стадия активности
Окончание стадии послелиночного доразвития проявляется в приобретении клещами способности к нападению на хозяев, т. е. в возникновении у них состояния агрессивности. В регионах с умеренным и субтропическим климатом стадии активности ограничены определенными сезонами года. Каждый из видов обладает свойственными ему сезонами активности имаго, нимф и личинок, которые могут перекрываться или бывают разделены во времени. Стадия активности обычно подразделяется на несколько периодов подстерегания, разделенных периодами покоя. У некоторых видов, и в частности у имаго представителей рода Dermacentor, отдельная особь может иметь весенний и осенний периоды подстерегания, разделенные летней поведенческой диапаузой, когда состояние агрессивности временно исчезает. Часть клещей, не сумевших найти прокормителей в первый сезон активности, зимует, и их активность возобновляется на второй и даже третий годы жизни.
У видов с подстерегающим типом нападения активизировавшиеся клещи поднимаются на растения или на поверхность растительной подстилки. На растениях, как живых, так и высохших, они в подавляющей массе занимают «позицию подстерегания» вблизи концов стеблей, листьев или сухих соломинок. Гнатосома подобных особей направлена вверх или вниз под разными углами к стволу или горизонтальной поверхности листа. II и III пары ног используются для удержания и обхватывают или зацепляются за субстрат. I пара ног у покоящихся особей слегка поджата к туловищу, а при раздражающих стимулах, исходящих от приближающегося хозяина, выдвинута вперед и совершает колебательные движения (рис. 64).

Высота размещения на растениях разных видов и фаз развития одного вида значительно варьирует (табл. 8). Местоположение активных клещей на растительности наиболее детально исследовано для I. ricinus (Gigon, 1985). В условиях мезофильного широколиственного леса клещи не обнаруживали четкой избирательности к определенным видам растений в подлеске. Основная масса взрослых располагалась в зоне от 2 до 80 см над поверхностью почвы, но единичные особи поднимались вверх до 170 см, а средняя высота составляла 45 см. Нимфы находились на высоте от 2 до 80 см (в среднем 30 см) и личинки — на высоте от 2 до 50 см.
В таежных лесах Дальнего Востока (рис. 65) взрослые I. persulcatus обнаруживались на высоте до 75 см (Померанцев, Сердюкова, 1948). Активные нимфы концентрировались на высоте 4—10 см, а личинки в лесной подстилке на глубине до 1 см и на растительности — на высоте до 5 см (Шлугер, 1961). В лесных местообитаниях I. scapularis в Миссисипи самцы и самки визуально обнаруживались на растительности на высоте от 40 до 87.5 см. При сборах на флаг 36 % клещей находилось на высоте 31—40 см, 18.6 % — 40—50 см и 21 % — 51— 60 см (Goddard, 1992).

Многие виды клещей подстерегают хозяев на растительности в открытых стациях. Клещи D. reticulatus в период их весенней активности на лугах и вырубках в средней полосе России обычно располагаются на высоте от нескольких сантиметров до одного метра от поверхности почвы, предпочитая сухие злаковые сопомины (Олсуфьев, 1953). По нашим наблюдениям, в условиях горной степи и полупустыни в Туркмении взрослые R. turanicus находились на растительности на высотах от 5 до 70 см, H. punctata — на 5—80 см и D. marginatus — на высоте 5—90 см.
Высота размещения клещей внутри травяно-кустарникового яруса определяется микроклиматическими факторами и видовой принадлежностью, и в ряде случаев доказана ее связь с высотой тела хозяина. Специальные исследования этого вопроса выполнены для немногих видов. Было установлено, что раз поднявшись на растительность, клещи не остаются на ней постоянно, но могут совершать периодические вертикальные миграции от вершин стеблей растений до их оснований. Впервые этот феномен был обнаружен для I. ricinus на пастбищах в Англии (Lees, Milne, 1951) и в дальнейшем детально исследован (Gigon, 1985) в широколиственном лесу в Швейцарии. В кустарниково-травяном ярусе, где находятся активные клещи этого вида, суточные и сезонные колебания влажности сглажены и во многом зависят от типа леса и густоты травяного покрова. В самом травяном ярусе существует гигротермический градиент. В верхней части яруса абсолютные показатели относительной влажности и температуры мало отличаются от макроклимата леса в целом. Напротив, ближе к поверхности растительной подстилки и внутри последней поддерживается наиболее высокая влажность (> 90 %) и наблюдаются наименьшие колебания температуры.
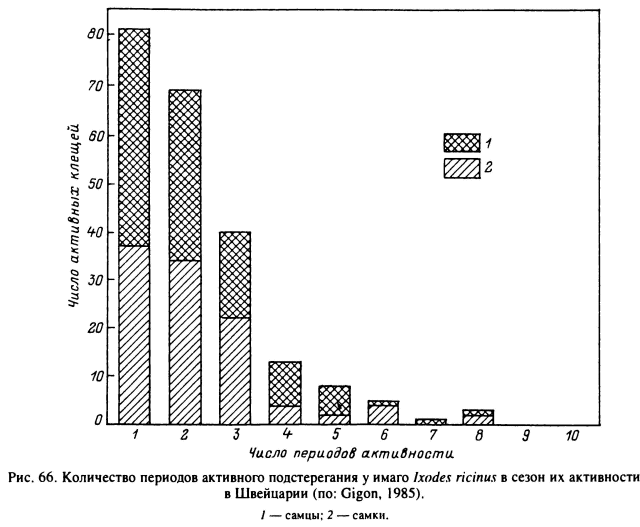
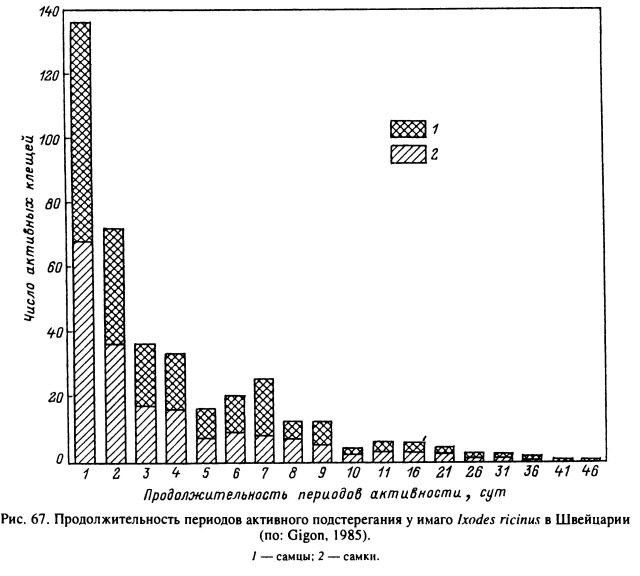
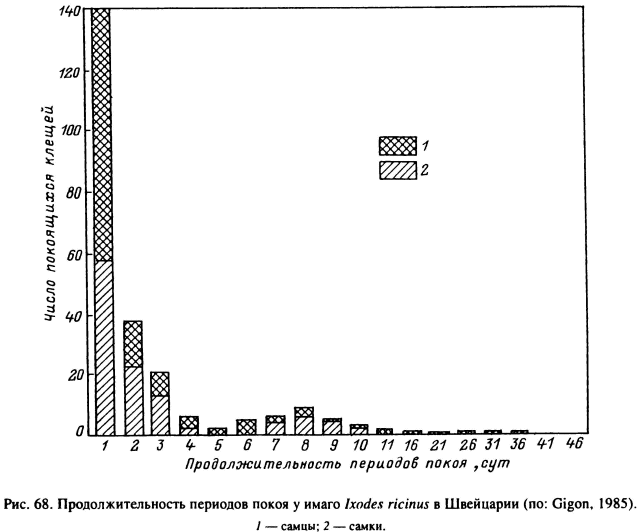
Даже во влажном микроклимате леса клещи I. ricinus при нахождении в верхней части травяного яруса испаряют воду и вынуждены периодически опускаться на поверхность и внутрь растительной подстилки, где и восполняют потери воды (Lees, Milne, 1951). Жизнь особи, подстерегающей хозяина, складывается из чередующихся фаз активности на растительности и покоя в подстилке. В условиях Швейцарии за индивидуальную жизнь голодной взрослой особи может быть от 1 до 8 фаз активности (рис. 66), продолжительность каждой из которых у большинства клещей варьирует от 1 до 9 сут (рис. 67), но в единичных случаях достигает 26—36 сут. Соответственно длительность фаз покоя варьирует от 1 до 4 сут, максимальная — до 36 сут. В среднем на время подстерегания в Швейцарии падает 75 % и на отдых — 25 % жизни голодных клещей (рис. 68). В Англии, где клещи живут на открытых пастбищах с более сухим микроклиматом, подстерегание занимает около 30 % и отдых — 70 % от времени жизни (Lees, Milne, 1951). Частота и продолжительность вертикальных миграций у этого вида в большей степени зависели от водного баланса организма и в меньшей степени от прямых воздействий температур и влажности окружающей среды. В Карелии взрослые I. ricinus в летние месяцы совершают регулярные вертикальные миграции вдоль стеблей растений, а осенью при понижении температуры и повышении относительной влажности воздуха в течение нескольких дней остаются на вершинах стеблей (Лутта, Шульман, 1958).
У клещей R. appendiculatus продолжительность периодов активного подстерегания на растительности и покоя в подстилке также определяется в первую очередь содержанием воды в их организме. Однако в условиях жаркого климата Кении с чередованием сезонов дождей и засухи температура и влажность окружающей среды уже в течение нескольких суток или даже часов могут нарушить водный баланс организма и вызвать прямую вертикальную миграцию как ответную реакцию на эти факторы (Punyua et al., 1985a, 1985b).
Непосредственная реакция на температуру и влажность наблюдается у голодных иксодид в аридных местообитаниях. Так, в Калифорнии в кустарниковых зарослях взрослые A. americanum в феврале, когда температура воздуха низкая, а влажность высокая, встречаются исключительно на поверхности почвы. Подъем их на растительность происходил в середине или конце апреля, и пик на растительности был зарегистрирован в конце мая—начале июня. С начала июля в связи с повышением температуры и уменьшением влажности воздуха клещи вновь перемещались в растительную подстилку и в почву, а с конца июля с началом снижения температуры и повышением влажности число клещей на растительности снова возрастало. Гидротермический режим определял и суточный ритм миграций. В мае количество клещей на растительности увеличивалось с утра к вечеру, в июне их численность была постоянной в течение всего дня, а в июле они спускались с растительности в наиболее жаркие дневные часы. В открытых стациях клещи были вынуждены проводить в почве и подстилке большее время, чем при наличии тени от деревьев (Semtner, Hair, 1973). Также и у клеща I. pacificus в условиях сухих лесов Калифорнии количество активных взрослых на растительности увеличивается с ростом влажности и с понижением температуры (Loye, Lane, 1988).
Еще большая зависимость вертикальной миграции от влажности обнаружена у личинок A. hebraeum, R. appendiculatus и R. evertsi в аридных открытых пастбищах в Южной Африке (Rechav, 1979). У них выявлен суточный ритм вертикальных миграций, так что наибольшее количество личинок на растительности наблюдалось с 6 до 9 и после 18 ч, когда влажность была максимальной, и наименьшее — в 12 ч.
В наибольшей степени сезонная и суточная активность зависит от температуры и влажности у взрослых H. asiaticum, которые не поднимаются на растительность, а подстерегают и активно преследуют хозяев на поверхности почвы. Этот вид успешно адаптировался к обитанию в жарких песчаных пустынях Средней Азии, где летом поверхность песка нагревается до 60—70 °С, а редкая растительность из эфемеров полностью выгорает. По нашим наблюдениям (Балашов, 1960), в начале апреля при дневных максимумах температуры не более 25 °С клещи бегают по поверхности песка и преследуют хозяев в течение всего дня. В конце апреля—начале мая, когда дневные температуры воздуха повышаются до 30 °С и более, поверхности почвы — до 45—50 °С, а относительная влажность воздуха понижается в полдень до 15—20 %, активность клещей наблюдается с рассвета до 9—10 ч и затем после 18 ч. В дневные часы клещи прячутся в норах песчанок и других животных и глубоких трещинах в почве, под корнями немногочисленных кустарников. В наиболее жаркие летние месяцы клещи бывают активны в поздневечерние, ночные и ранние утренние часы. Наряду с суточным ритмом активности у H. asiaticum обнаружены чередования периодов активности на поверхности на протяжении 10—20 сут и 2—4-суточных периодов покоя в убежищах с высокой относительной влажностью.
Освещенность и смена дня и ночи влияют на суточный ритм активности в значительно меньшей степени, чем температура и влажность. Для I. ricinus (Lees, Milne, 1951; Gigon, 1985), I. scapularis (Durden et al., 1996), D. occidentalis (Lane et al., 1985) и немногих других изученных видов выявлена круглосуточная активность подстерегания и вертикальных миграций. Недостаток данных по этому вопросу связан с техническими трудностями учетов активности клещей в темноте.
В Красноярском крае у взрослых I. persulcatus в теплую солнечную погоду наблюдаются утренний и вечерний пики активности при значительном снижении в дневные часы. В дни с пасмурной погодой отмечена одновершинная кривая активности с минимумом ранним утром, высокой активностью днем и пиком в послеполуденные часы (Бабенко, Хижинский, 1964). Для нимф таежного клеща характерен пик активности в конце вечерних сумерок со спадом при наступлении ночной темноты (Бабенко, 1974).
У голодных взрослых D. marginatus на открытых горных пастбищах в Армении в период осенней активности в теплую погоду наибольшее количество подстерегающих клещей на стеблях растений отмечено вечером (74 %) и значительно меньше утром (16 %) и днем (7.5 %) (Рухкян, 1987). У голодных D. variabilis активность подъема на растительность личинок и имаго находилась в прямой зависимости от интенсивности солнечного освещения. В пасмурную погоду она сильно снижается даже при достаточно высоких температурах (Atwood, Sonenshine, 1967). У обитающих в тени под пологом леса I. persulcatus и Н. concinna мы, напротив, наблюдали активное перемещение клещей на затененную сторону стебля или листа при прямом солнечном освещении в дневные часы.
Различную высоту размещения клещей на растительности некоторые исследователи связывают с приближением их к зонам активных передвижений главных хозяев. По наблюдениям М. Н. Слонова (1963), в южнотаежных лесах Дальнего Востока наблюдаются значительные различия в ярусах максимальной концентрации активных личинок, нимф и взрослых I. persulcatus, I. pomeranzevi, H. concinna, H. japonica, D. silvarum. Этим объясняется разная пораженность местных видов млекопитающих, передвигающихся в разных ярусах леса. На кротах и бурозубках, перемещающихся только в подстилке или по ее поверхности, паразитируют исключительно личинки I. persulcatus, а на ежах, перемещающихся только по поверхности подстилки в нижнем травяном ярусе, — личинки и нимфы всех 5 видов. Равным образом личинками и нимфами этих клещей были поражены белки и бурундуки, активные в нескольких растительных ярусах.
Имаго клещей, нападающих на крупных и средних млекопитающих, находятся на растительности в зоне от 30 до 60 см от поверхности почвы (табл. 8), что обеспечивает их контакт с шерстью живота и ног. Высота размещения на растительности нимф и особенно личинок, питающихся на мелких и средних млекопитающих, обычно значительно меньше, чем у имаго. Исключение составляют личинки Boophilus, размещающиеся на высоте до 50 см и более, что соответствует длине ног хозяина — крупного рогатого скота. Наибольшая высота подъема на растительность отмечена у личинок D. albipictus в Северной Америке. Главным хозяином для них служит лось, и они подстерегают его на высоте от 20 до 220 см от поверхности земли (Wilkinson et al., 1982). В Калифорнии подстерегающие нимфы D. occidentalis находятся непосредственно на поверхности почвы или не выше 4—10 см от ее поверхности на растительности. Это совпадает с зоной активности их прокормителей — грызунов и зайцеобразных. В тех же местообитаниях нимфы I. pacificus вообще не поднимаются на растительность и подстерегают своих прокормителей — ящериц на поверхности или внутри растительного опада (Lane et al., 1995).
Скорость перемещений клещей по горизонтали может быть достаточно велика. По наблюдениям Н. М. Окуловой (1986), в естественных местообитаниях самки D. silvarum перемещались со скоростью до 36.4 см/мин, D. reticulatus — 39.2, I. persulcatus — 15.3—23.8, I. ricinus — 12.8, H. concinna — 20.8, I. trianguliceps — 3.6 и I. pomeranzevi — 5.5 см/мин. По нашим наблюдениям, при температуре воздуха 24—26 °С голодные взрослые особи Н. asiaticum пробегали в направлении хозяина до 100 и более сантиметров в минуту. Скорость передвижения клещей зависит от размеров их тела, так что у взрослых она больше, чем у личинок и нимф. Как биологический процесс, она определяется температурой и достигает максимальных значений при приближении к верхнему порогу активности. При отсутствии привлекающих стимулов эти передвижения беспорядочны и непродолжительны. У большинства видов иксодид с подстерегающим типом нападения способность голодных клещей к активным горизонтальным перемещениям внутри занимаемых ими местообитаний ограничена несколькими метрами движения у взрослых и сантиметрами у личинок (табл. 9). Эти результаты получены путем выпуска меченых клещей и последующего их отлова на флаг или же на приманку в виде источника СО2.
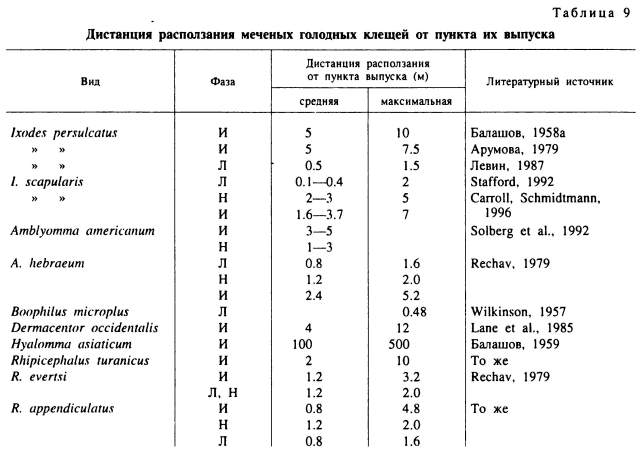
У I. persulcatus в различных типах таежных лесов самцы и самки за время от нескольких дней до 1 мес отползали от места выпуска не далее 10 м, а основная их масса оставалась в радиусе 1—2 м (Балашов, 1958а; Арумова, 1979). Личинки в тех же условиях оставались в радиусе до 0.5 м, и только единичные особи отползали на 1.5 м (Левин, 1987). Столь же ограничены способности к горизонтальным миграциям у I. ricinus (Lees, Milne, 1951; Gigon, 1985). У I. seapularis большая часть личинок оставалась в радиусе 0—30 см от места вылупления (Stafford, 1992), меченые нимфы расползались на расстояние до 2—3 м и взрослые особи — до 5 м (Carroll, Schmidtmann, 1996). При сборах ловушкой с СО2 меченые самцы и самки I. scapularis в основной массе были собраны в радиусе до 1.8 м и только единичные с расстояния в 5 м (Falco, Fish, 1991). У более крупного по размерам тела Д. americanum нимфы и взрослые отлавливались источником СО2 с расстояния до 9.3 м, а личинки — 1.5 м (Koch, McNew, 1982).
Ограниченная подвижность свойственна и видам из открытых местообитаний. В Туркмении в фисташковом редколесье до 80 % меченых взрослых R. turanicus были отловлены на расстояниях до 1—2 м от места выпуска и только немногие в 5—10 м (Балашов, 1959). Взрослых D. occidentalis собирали на СО2 в кустарниковых зарослях с расстояний до 12 м, но основную массу с 0.5—4 м (Lane et al., 1985). На пастбищах в ЮАР взрослые A. hebraeum были отловлены на расстояниях в 0.4—2.4 м от места выпуска, R. appendiculatus и R. evertsi — 0.4—0.8 м и только немногие на расстоянии до 5 м. Личинки этих видов оставались на расстоянии до 1 м (Rechav, 1979). Исключение составляет Н. asiaticum и, вероятно, другие виды этого рода, активно преследующие своих хозяев. В песчаной пустыне в Туркмении меченые взрослые особи были отловлены через 5—30 сут на расстояниях до 500 м от места выпуска. Однако наибольшее количество клещей оставалось в радиусе 25—100 м (Балашов, 1959).
В связи с весьма ограниченной способностью к активным перемещениям голодные стадии клещей не имеют существенного значения в их расселении. Однако для личинок В. microplus установлена возможность их пассивного переноса ветром на расстояния до 30 м, а в отдельных случаях и до 80 м (Lewis, 1970), тогда как самостоятельно они преодолевают расстояние не более 40 см (Wilkinson, 1957). Главная роль в расселении иксодид принадлежит их хозяевам, которые в зависимости от их миграционных способностей разносят напитавшихся клещей на расстояния от нескольких сотен метров до десятков и сотен километров от места нападения голодных особей.