Иксодовые клещи — паразиты и переносчики инфекций
Прикрепление
С момента прицепления клеща к хозяину и до начала прикрепления с помощью ротовых органов к его коже паразит удерживается в шерсти и на поверхности кожи благодаря разного рода фиксаторным крючьям, шипам и щетинкам. Важная роль в этом принадлежит ногам, которые не только обеспечивают закрепление, но и служат для активного перемещения по телу хозяина. Они же удерживают паразита при отряхиваниях и других механических оборонительных реакциях хозяина. Как и у других эктопаразитов, здесь особенно важны парные коготки на лапках. Они одинаково хорошо развиты как у быстро бегающих видов рода Нуalomma, так и у пассивно ожидающих клещей родов Ixodes, Dermacentor или Rhipicephalus, у паразитов млекопитающих и рептилий (табл. XIII, 1; XXIX, 1).
Фиксация клеща среди волос может обеспечиваться направленными назад кутикулярными выростами основания головки (аурикулы, дорсальные и вентральные корнуа) и зубцевидными выростами на коксах и вертлугах ног (табл. XIII, 2—4). При подгибании основания гнатосомы или ограниченных смещениях кокс волосы оказываются зажатыми между соответствующими кутикулярными выростами и идиосомой клеща. Некоторыми видами Ixodes и Haemaphysalis волосы зажимаются между выростами I членика пальп и передним выступом кокс ног I пары при дорсовентральном перемещении гнатосомы.
Предполагают, что форма и размеры гнатосомальных и коксальных фиксаторных выростов в какой-то мере соответствуют особенностям волосяного покрова хозяев. Так, среди нескольких видов рода Haemaphysalis, паразитирующих на тенреках, у самцов и самок клещей, прикрепляющихся на ушных раковинах с почти голой кожей, коксальные зубцы развиты слабо. У видов же, присасывающихся среди густого волосяного покрова и жестких игл туловища, коксальные зубцы отличаются особенно мощным развитием. В роде Amblyomma для многих паразитов млекопитающих характерно сильное развитие каудальных шипов кокс IV, а у паразитов рептилий они отсутствуют. Не развиты коксальные шипы у представителей рода Аропотта — облигатных паразитов рептилий (Померанцев, 1948а; Hoogstraal, Kim, 1985).
После активного или пассивного зацепления с помощью ног за тело хозяина клещ некоторое время активно перемещается по поверхности тела, прежде чем он выберет характерное для вида место прикрепления. Выбор места прикрепления на теле хозяина осуществляется активно, так что в случае нападения на крупных копытных места первичного зацепления паразита за ноги и брюхо хозяина и окончательного присасывания могут быть разделены значительным расстоянием. Направление движения к месту прикрепления, по-видимому, определяется какими-то стимулами организма прокормителя или градиентом микроклиматических факторов (температура, освещенность, гравитация и др.), но, к сожалению, эти факторы остаются неисследованными.
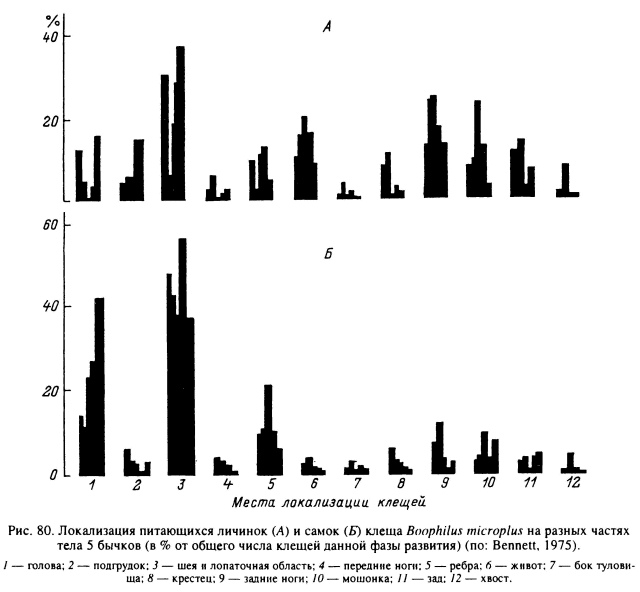
Голодные имаго клещей, подстерегающие копытных животных на растительности, обычно сперва зацепляются за передние ноги, голову или шею прокормителя, после чего переползают на свои видоспецифичные местообитания. Подобная же миграция отмечена и у личинок В. microplus (рис. 80), первоначально зацепляющихся за передние ноги и дистальную часть головы крупного рогатого скота, откуда они переползают в направлении шеи, лопаток и ушей (Bennett, 1975). Активная миграция к местам присасывания была показана для личинок I. ricinus. Последние независимо от места первоначального прикрепления к телу грызуна сразу же занимают позицию «против шерсти» и затем перемещаются в шерстном покрове или по его поверхности в сторону головы, где и присасываются (Nilsson, Lundqvist, 1978).
Успешное питание клеща возможно, если он сможет прорезать наружный роговой слой кожи, достаточно прочно и на длительный срок закрепиться и иметь возможность беспрепятственно отсасывать из толщи кожи хозяина кровь, воспалительный инфильтрат и продукты лизиса тканей. Прорезание кожи хозяина и последующее удержание в течение многих дней в образовавшейся в месте прикрепления ранке обеспечивается как высокоспециализированным ротовым аппаратом, так и уникальным приклеиванием клеща к коже секретом слюнных желез. Чисто визуально, по соотношению длин хоботка и основания гнатосомы у имагинальной фазы, иксодид разделяют на длиннохоботковые (Amblyomma, Аропотта, Hyalomma, большая часть lxodes) и короткохоботковые виды (Haemaphysalis, Dermacentor, Boophilus, Rhipicephalus). Подобная классификация, несмотря на многочисленные исключения на неполовозрелых фазах, имеет под собой физиологическую основу, связанную со способом удержания клещей в коже хозяина.
Мощное вооружение пальцев хелицер (табл. II, 4; XXIX, 3) позволяет с помощью их попеременных движений прорезать отверстие в роговом слое эпидермиса за первые 5—20 мин прикрепления. Толщина рогового слоя в местах питания иксодид может варьировать от 5—15 мкм у мелких млекопитающих до 20— 50 мкм и более у крупных копытных. Несмотря на высокую механическую прочность этого слоя, он, по-видимому, не создает серьезного барьера на пути ротовых частей клеща к внутренним слоям кожи а кровеносными сосудами. При этом не обнаруживается прямой связи между размерами и вооружением пальцев хелицер и толщиной рогового слоя. Во всяком случае, на крупном рогатом скоте и диких копытных наряду со взрослыми клещами успешно питаются и личинки некоторых видов иксодид, включая весь род Boophilus.
Среди Ixodinae известны виды, у которых цементный футляр отсутствует, как например у самок I. holocyclus (Stone, Binnington, 1989), I. ricinus (Павловский, Алфеева, 1941; Brossard, Fivaz, 1982), I. persulcatus (наши данные). С начала прикрепления и до отпадения напитавшейся особи хоботок этих видов клещей полностью погружен в кожу хозяина и непосредственно контактирует с окружающими его элементами дермы и клетками воспалительного инфильтрата (рис. 77; табл. XXXVII, 3). Среди Ixodinae цементный футляр был описан у I. tasmani (Moorhouse, 1969), нимф I. urae (Eveleigh et al., 1952) и личинок I. ricinus (Амосова, 1994), но во всех этих случаях он развит значительно слабее, чем у Amblyomminae.
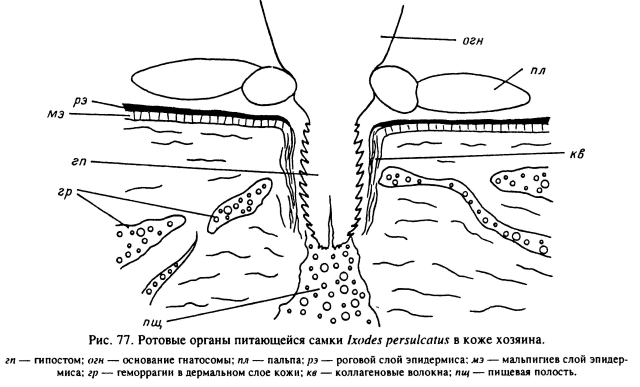
У длинно- и короткохоботковых видов Amblyomminae после первоначального прорезания рогового слоя кожи механизм прикрепления различен. У относящихся к первой группе представителей родов Hyalomma, Amblyomma, Aponomma (рис. 78) в ранку погружается весь хоботок, практически до основания головки, а пальпы отгибаются почти параллельно поверхности кожи. В результате длина гнатосомы у этих клещей довольно точно отражает глубину проникновения ротовых частей внутрь кожи (табл. 15). У личинок и нимф концы выдвинутых вперед пальцев хелицер и гипостома обычно находятся в богатом кровеносными сосудами сосочковом слое дермы или на границе ее с мальпигиевым слоем эпидермиса. У взрослых особей, особенно у крупных видов из родов Hyalomma и Amblyomma, при длине гипостома 0.8—0.9 мм хоботок может пронизывать всю толщу кожи, вплоть до подкожной клетчатки (Балашов, 1965; Brown, Knapp, 1980a, 1980b). Непосредственно от тканей хозяина хоботок этих клещей отделен застывшим секретом слюнных желез — цементным футляром (табл. XXXVII, 1, 2). Последний имеет форму трубки и заходит в кожу хозяина несколько дальше дистальных окончаний хелицер и гипостома. Соответственно пища попадает сначала во внутренний канал дистального конца футляра и из него уже в предротовую полость клеща. На поверхности кожи вокруг основания хоботка застывший слюнной секрет образует лишь небольшой плоский ободок, являющийся продолжением цементного футляра. Последний вклеивается своими концентрическими наплывами и выростами непосредственно в пространство между клетками и неклеточными волокнами дермы и эпидермиса. Сам клещ прочно удерживается внутри футляра мощными зубцами гипостома и более мелкими зубчиками поверхности наружных хелицеральных футляров. Эти кутикулярные элементы образуют свои отпечатки в застывающей слюне (табл. XXXVII, 2), и хоботок оказывается как бы заключенным внутри цементного футляра, образуя с ним единую структуру.
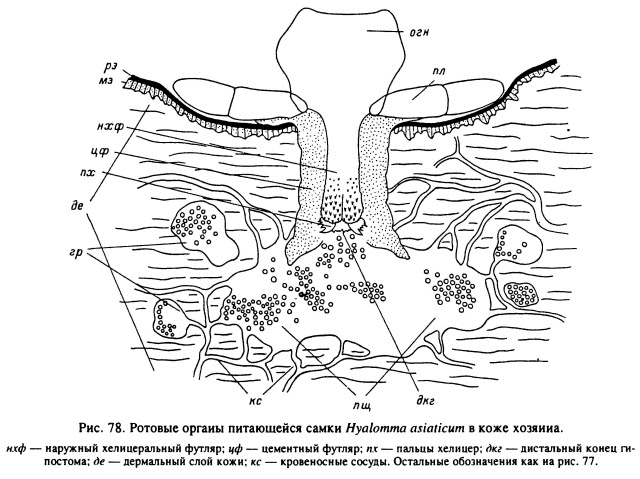
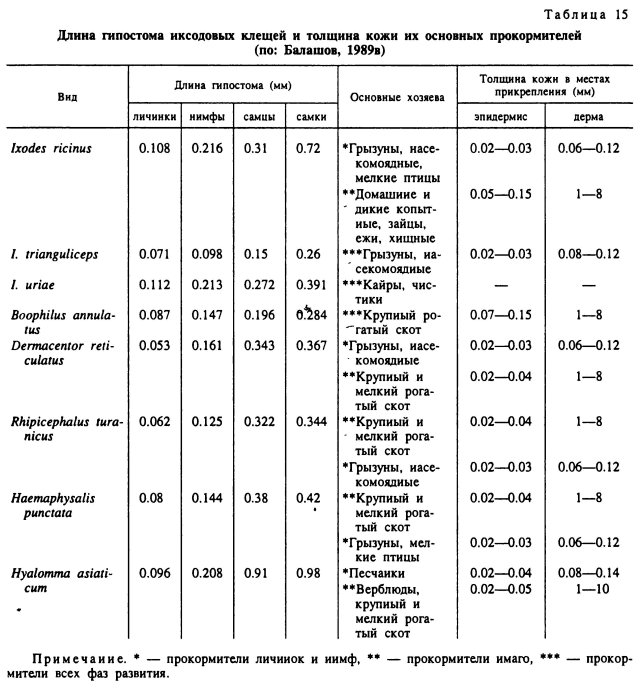
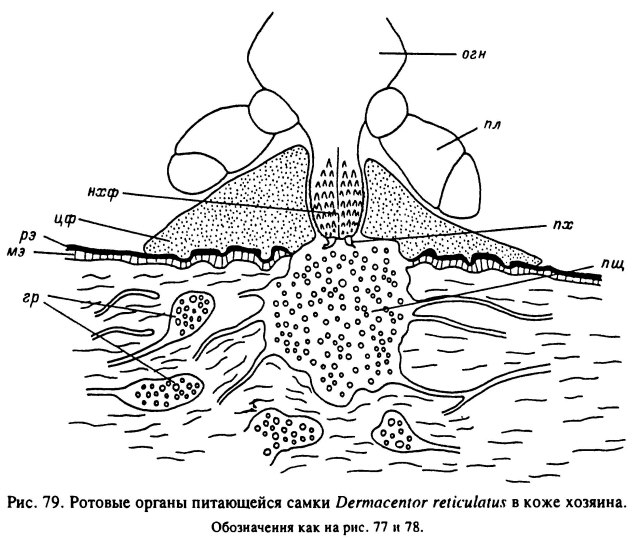
У клещей D. marginatus (Амосова, 1989), R. sanguineus (Theis, Budwiser, 1974), В. microplus (Moorhouse, Tatchell, 1966; Tatchell, Moorhouse, 1968) и нескольких представителей Haemaphysalis (Moorhouse, 1969; Chinery, 1973), относимых к группе короткохоботковых видов (рис. 79), большая часть хоботка питающегося клеща остается над поверхностью кожи прокормителя. При этом клещ как бы приклеивается к поверхности кожи конусовидным цементным футляром, плотно прилегающим к поверхности гипостома и наружных хелицеральных футляров вплоть до основания гнатосомы. Слегка раздвинутые пальпы при этом остаются снаружи от конуса цементного футляра. Расширенное основание последнего образует многочисленные натеки, заходящие в межклеточные щели рогового и зернистого слоев эпидермиса. В кожу хозяина сквозь ранку входят только выдвинутые в переднее положение пальцы хелицер, не заходящие глубже границы мальпигиевого слоя с дермой. Внутренняя часть цементного футляра развита значительно слабее наружной. Она также не заходит далее мальпигиевого слоя и окружает снаружи пальцы хелицер. Общая длина хоботка, несмотря на большие различия у разных видов и фаз развития одного вида, существенно не влияет на глубину погружения ротовых органов в кожу хозяина. Например, по нашим данным, длина гипостома увеличивается у В. annulatus с 0.08—0.09 мм у личинок до 0.2—0.3 мм у самок, но пальцы хелицер питающихся особей остаются в мальпигиевом слое эпидермиса на глубине до 0.2—0.3 мм от поверхности кожи (табл. 15).
Сопоставление длины хоботков и толщины кожи главных прокормителей иксодид в местах прикрепления этих клещей во многих случаях не выявляет положительной связи между этими параметрами (табл. 15). Закрепление ротовых органов представителей родов Boophilus, Rhipicephalus, Dermacentor с помощью цементного футляра над поверхностью кожи и отсасывание пищи из очага воспаления в толще дермы под местом прикрепления паразита обеспечивает нормальное питание не только имаго, но и мелких преимагинальных особей на копытных с толстой кожей.
Многие виды Ixodes на всех фазах развития, a Hyalomma и Amblyomma на неполовозрелых фазах, несмотря на длинный хоботок, успешно прокармливаются на животных с наиболее тонкой кожей — грызунах и насекомоядных. Ограничения в выборе хозяев у иксодид связаны, скорее, не с толщиной кожи, а с размерами тела млекопитающего. Виды клещей, питающиеся на имагинальной фазе на мелких животных, обладают и более мелкими размерами тела (I. trianguliceps, I. subterraneus, I. occultus, R. schulzei). Напротив, паразиты особенно крупных млекопитающих, как например Н. asiaticum и Н. dromedarii с верблюдов или A. tholloni со слонов, принадлежат к наиболее крупным видам клещей. Подобная закономерность обусловлена многими факторами, включая чрезмерное разрушение покровов и интоксикацию мелкого хозяина крупным паразитом и невозможностью завершения многодневного питания до гибели хозяина.
Поверхностное прикрепление клещей к коже, по-видимому, дает паразиту определенные преимущества, особенно в случае с двуххозяинными и однохозяинными видами. Меньшее травмирование кожи в месте прикрепления, encephalitis.ru вероятно, снижает и уровень ответных защитных реакций на протяжении непрерывного питания личинок, нимф и имаго клещей на одном и том же участке тела. Интересно, что эти же короткохоботковые одно- и двуххозяинные клещи отличаются и более узкой пищевой специализацией по сравнению с некоторыми длиннохоботковыми треххозяинными видами. К числу последних, в частности, принадлежат I. persulcatus и А. americanum, являющиеся рекордсменами по числу видов хозяев. Правда, среди длинно-хоботковых видов рода Hyalomma известны двуххозяинные и однохозяинные паразиты копытных, так что роль коротко- и длиннохоботковости в эволюции паразито-хозяинных отношений иксодид требует дальнейших исследований.