Иксодовые клещи — паразиты и переносчики инфекций
Места прикрепления
Для большинства видов иксодид характерно существование мест их преимущественного прикрепления на теле хозяев, тогда как на других частях тела они встречаются значительно реже или вообще отсутствуют. Наиболее детально изучены места прикрепления клещей на теле сельскохозяйственных животных.
В условиях Северо-Запада России и в Западной Сибири 60—70 % самок I. persulcatus присасываются на шее и подгрудке крупного рогатого скота, 20—24 % — в паху и на голове, а в прочих местах не более 10 % (Таежный клещ..., 1985). Самки I. ricinus присасываются преимущественно в паховой области и на вымени, D. reticulatus — на голове и шее крупного рогатого скота (табл. 16). В Средней Азии клещи рода Rhipicephalus избегают участков тела овцы, которые плотно покрыты густой шерстью, пропитанной жиром и потом. Соответственно имаго R. turanicus прикрепляются главным образом на ушах, в паху и околоанальной области. R. bursa паразитируют на вымени, в паху и у основания хвоста. После стрижки овец эти же виды присасываются к шее, спине и бокам животных (Балашов, 1982).
В Южной Африке (Howell et al., 1978) личинки и нимфы R. evertsi прикрепляются преимущественно на внутренней стороне ушей у крупного рогатого скота, а имаго — под основанием хвоста и в паху. На овцах и козах этот вид прикрепляется на груди и в подмышечных областях. У R. appendiculatus при слабой пораженности скота клещи питаются главным образом на ушах, а при большой пораженности встречаются по всей голове и на других участках тела. Нимфы прикрепляются на ушах и вокруг их оснований, на шее и в меньшем количестве на туловище и ногах. На собаках взрослые R. sanguineus предпочитают уши и шею, но при сильном поражении встречаются на других участках тела. Повсеместно прикрепляются личинки и нимфы этого вида. Н. marginatum rufipes питаются в околоанальной области, на гениталиях и редко — на ногах. У A. hebraeum взрослые выбирают на скоте нижнюю часть туловища и особенно лишенные волос пах, подмышки, подгрудок и живот. Нимфы прикрепляются на подгрудке, груди и ногах, личинки — на голове. На овцах взрослые особи этого вида прикрепляются в паху, подмышках, на груди и в околоанальной зоне. У A. variegatum места прикрепления те же, что и у A. hebraeum. У взрослых I. rubicundus (Fourie, Zyl, 1991) при паразитировании на овцах и козах большая часть клещей питается на передней половине тела этих животных, а на антилопах — также и на задних ногах.
У однохозяинного В. microplus при паразитировании на крупном рогатом скоте личинки локализуются преимущественно на голове, шее и лопатках, причем количество их резко снижается в направлении задней половины туловища. На голове большинство личинок находится на ушах по их внутреннему краю, а на шее и лопатках — вдоль хребта. Локализация нимф сходна с таковой у личинок, но более равномерна по всему телу. Самки и самцы в равной мере питаются и на передней, и на задней половинах тела, где они особенно многочисленны в паховой области и на внутренней стороне бедер (рис. 80) (Bennett, 1975).
У A. americanum более 50 % личинок питаются на ушах крупного рогатого скота, а нимфы кроме головы и ушей также на груди, передних ногах и шее. Более 90 % взрослых клещей прикрепляются в подмышечной, паховой и околоанальной областях тела, с боков туловища, на вымени (Barnard et al., 1989). На оленях самки и самцы этого вида прикрепляются главным образом с наружной стороны ушей, на голове, вымени, а нимфы и личинки — с внутренней стороны ушей. Только на ушах питалось 54 % всех самок, 67 % самцов, 97 % нимф и 81 % личинок (Bloemer et al., 1988).
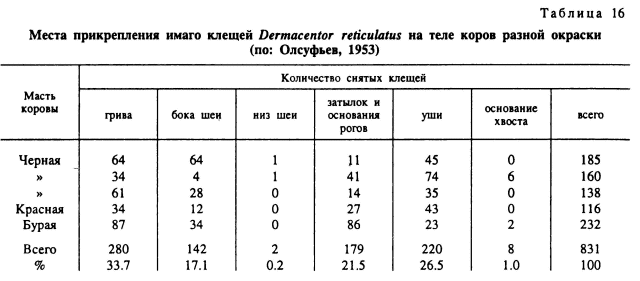
На млекопитающих со средними размерами тела клещи прикрепляются преимущественно к голове, шее и значительно реже — к туловищу и ногам. При одновременном паразитировании на диком кролике (Oryctolagus cuniculi) I. ventalloi, H. hispanica и R. pusillus отмечено предпочтение определенных частей головы для каждого из этих видов (рис. 81) (Marquez, Guiguen, 1992). R. pumilio, паразитирующий на всех фазах на зайцах-толаях (Lepustolai), прикрепляется в основном на ушах (Гребенюк, 1966). Исключение составляют ежи, на которых различные виды клещей распределены по всему телу более равномерно (Кучерук и др., 1956).
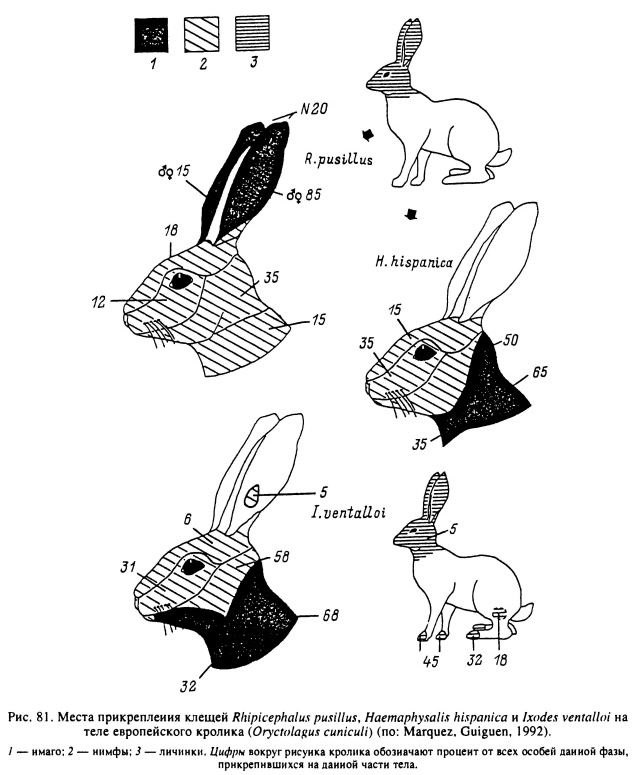
У грызунов и насекомоядных иксодовые клещи питаются главным образом на ушах, других частях головы и реже на туловище. Так, при осмотре на заклещевленность личинками I. persulcatus 4167 мелких млекопитающих 20 видов 75 % клещей обнаружено на ушах, 24 — на голове и 3 % — на туловище, при определенных отклонениях этих показателей для разных видов прокормителей. Нимфы клеща распределялись на тех же видах хозяев в следующей пропорции: уши — 29—40 %, мордочка — 5—29, лоб — 6.5—40, щеки — 7—40 % (Васильева, 1964). Преимущественное прикрепление к голове и ушам грызунов было свойственно личинкам и нимфам D. variabilis, питавшимся на хлопковых крысах (Sigmodon hispidus) и оленьих мышах (Peromyscus maniculatus) в Северной Америке (табл. 17) (Gage et al., 1992b).
Определенная избирательность мест прикрепления клещей отмечена и у рептилий. На ящерицах различные виды родов Аропотта, Amblyomma, Ixodes, Haemaphysalis в наибольшем количестве присасываются на голове вокруг ушных отверстий, глаз, в углах рта и у оснований передних, реже задних ног, на спине. Ротовые части паразитов вводятся в участки с тонкой кожей под или между чешуями и щитками, в ушные раксяжны, веки (Andrews et al., 1982; Hesse, 1985; Таежный клещ..., 1985). Например, при нашем обследовании на заклещевленность 14 горных агам (Laudakia caucasia) в горах Кугитанга (Туркмения) из 298 питавшихся нимф Н. sulcata 201 была прикреплена вокруг шеи, 77 — у основания передних ног и только 11 — в других местах. Виды, паразитирующие на наземных черепахах, как например H. aegyptium, прикрепляются вокруг оснований задних ног и хвоста, реже — на шее и лишь в редких случаях — на поврежденных участках панциря (Petney, Al-Yaman, 1985; наши данные).
При нападении клещей на человека, несмотря на обувь и одежду, ограничивающие распространение этих паразитов по телу, также выявлены преимущественные места присасывания. По данным учета мест прикрепления самок I. persulcatus у 2500 жителей Карелии, наибольший процент присосавшихся клещей отмечен в подмышечной области и далее по степени убывания — на груди, животе, руках, ягодицах и ногах. У детей большое количество прикреплялось также к голове. Предполагают, что локализация присасываний попавших под одежду клещей в основном определяется сочетанием воздействий на соответствующие рецепторы паразита температуры, запахов, концентрации СО2, влажности прикожного слоя воздуха на разных частях поверхности тела (Розенберг, 1984).
Для объяснения приуроченности питающихся клещей к определенным частям тела хозяев предложено несколько гипотез. Не вызывает сомнений исключительное значение способности животных к самоочищению от паразитов с помощью оборонительных поведенческих реакций. К числу их относят стряхивание, слизывание, выгрызание, выклевывание и раздавливание паразитов. Сложный комплекс подобных реакций против клеща D. albipictus был описан у лосей (рис. 82) (Samuel, 1991). В результате питающиеся клещи концентрируются на частях тела, наименее доступных для механического самоочищения. Среди крупного и мелкого рогатого скота активные оборонительные реакции в значительной степени ограничивают места питания наиболее уязвимых взрослых особей многих видов клещей головой, подмышечной, паховой и околоанальной областями (Балашов, 1967; Barnard et al., 1989; Fourie, Zyl, 1991).
У грызунов, зайцеобразных и насекомоядных клещи питаются главным образом на ушах и других частях головы, реже — на шее, груди, под мышками и на туловище. В этих местах длительно питающиеся и неспособные менять место прикрепления клещи наименее доступны для вычесывания или выкусывания хозяином. Напротив, на ежах клещи защищены от оборонительных реакций хозяина иглами и распределены по его телу более равномерно (Кучерук и др., 1956; Nilsson, 1981). Исключительной способностью обезьян к индивидуальному и групповому самоочищению от паразитов можно объяснить не только их слабую пораженность, но и отсутствие у них специфических видов иксодид.
На птицах клещи присасываются в подавляющей массе на голове и верхней части шеи, причем у взрослых особей и птенцов их локализация может существенно отличаться. Локализация питающихся клещей на этих хозяевах во многом определяется высокой подвижностью их шеи и способностью выклевывать паразитов с большей части туловища и крыльев. На взрослых птицах и оперившихся птенцах клещи A. marginatum, A. punctata, I. lividus, I. subterraneus чаще всего прикрепляются вокруг глаз, основания клюва, ушей, на шее и реже — на поверхности части тела, особенно под крыльями. На голых и пуховых птенцах с меньшей способностью к самоочищению гнездово-норовые виды I. lividus и I. subterraneus присасываются к туловищу и особенно часто под крыльями. Несмотря на приведенные факты, все многообразие распределения клещей по телу хозяев трудно объяснить только различной способностью к самоочищению от паразитов.
Несомненно, что первостепенное значение для видоспецифической приуроченности мест питания клещей к определенным частям тела прокормителей имеет строение их кожи. К сожалению, если исключить бесспорные случаи невозможности питания клещей на чрезмерно ороговевших участках покровов (щитки, чешуи, мозоли и др.), а также в местах с очень густой шерстью, обильно пропитанной потом, то почти постоянное отсутствие или преимущественное питание этих паразитов на разных частях тела хозяина далеко не всегда поддается объяснению.
Важную роль в выборе местообитаний играет для клещей их микроклимат. Даже в норме разные участки поверхности тела млекопитающих имеют разную температуру (Murray, 1986). Еще большие различия в этом отношении между представителями разных отрядов млекопитающих, между млекопитающими и птицами. Поверхностное местоположение питающихся клещей делает их подверженными прямому воздействию факторов внешней среды, так что в аридных регионах с жарким и сухим климатом важную роль в определении температуры поверхности и прикожного слоя воздуха может играть инсоляция. По наблюдениям И. Г. Галузо (1947), в Средней Азии в летние месяцы у крупного рогатого скота, выпасающегося на открытых пастбищах, уши, спина и бока тела нагревались в полдень до 42— 43 °С, а на брюхе температура не превышала 38—39 °С. При этом части тела, покрытые белой шерстью, нагревались до 39 °С, а черной и бурой — до 43 °С. Взрослые особи A. detritum в подобных условиях встречались в основном на нижней половине туловища, защищенной от прямого солнечного воздействия, и отсутствовали на открытых частях тела.
В Австралии клещ В. microplus отсутствует на буйволах, отличающихся редкими волосами и темным цветом кожи, но при содержании этих животных в загонах, защищенных от солнца, они бывают в сильной степени поражены клещами (Tatchell, 1986). При паразитировании клещей этого вида на крупном рогатом скоте было показано, что личинки в большем проценте случаев развивались на животных, содержавшихся днем в тени и имевших более прохладную поверхность кожи, чем в случае со скотом, весь день находившимся на солнце и имевшим более высокую температуру кожи (О'Kelly, Spiers, 1983). Возможно, что преимущественное прикрепление взрослых клещей D. andersoni прерийной расы на нижней стороне туловища крупного рогатого скота, а клещей лесной расы — на верхней стороне также обусловлено выбором более затененных местообитаний в условиях более жаркого климата прерий (Wilkinson, 1972). В Африке питающиеся на слонах клещи A. tholloni были обнаружены наряду с областью паха и головы также в ротовой полости этих хозяев, а также в ноздрях гиппопотамов (Norval, 1983). Эти случаи, как и нахождение питающихся в ноздрях верблюдов A. dromedarii, объясняют активным выбором местообитаний с достаточно высокой влажностью и защитой от прямого солнца (Tatchell, 1986).
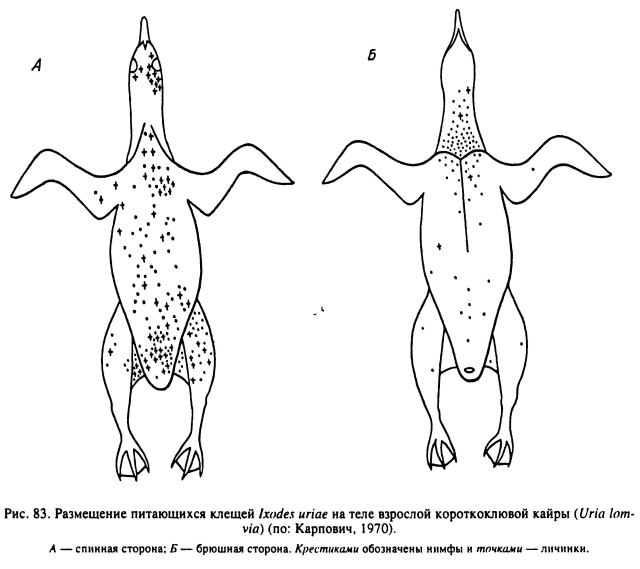
У паразитирующего на морских птицах I. uriae личинки и нимфы прикрепляются почти по всему телу хозяина — толстоклювой кайры (Uria lomvia). Нет их только на ногах и крыльях взрослых птиц (рис. 83). В то же время у птенцов различных видов чистиковых эти клещи, включая имаго, часто питаются на перепонке и цевке лапок, несмотря на низкую температуру их поверхности в 20 °С по сравнению с 40 °С на туловище. Предполагают (Карпович, 1970), что у морских птиц различия в местах прикрепления клещей на птенцах и взрослых особях наряду с температурой поверхности тела зависят также от степени контакта разных частей тела с холодной морской водой.
У клещей Н. aegiptium при паразитировании на средиземноморских черепахах (Testudo graeca) (Petney, Al-Yaman, 1985) и 3 видов Amblyomma и Аропотта на ящерицах сцинках в Австралии (Andrews, Petney, 1981) распределение питающихся клещей на теле хозяев зависит от плотности популяций паразитов. Снижение избирательности в местах прикрепления отмечено и при высокой пораженности сельскохозяйственных животных многими видами иксодид.
Для немногих видов иксодид при достаточно высокой их численности, когда на хозяине питается несколько особей, последние образуют характерные местные скопления. Клещи в этих случаях присасываются рядом с уже питающимися особями, так что на теле прокормителя возникают группы из нескольких паразитов. Иногда, как например в случае со взрослыми Н. asiaticum на верблюдах, Н. апаtolicum и Н. detritum на крупном рогатом скоте или R. turanicus на ушах овец, подобные агрегации включают до 30 и более питающихся самцов и самок от разных сроков прикрепления. У однохозяинных В. annulatus или Н. scupense по-
добные скопления включают личинок, нимф и взрослых, а у двуххозяинных Н. marginatum или R. bursa — личинок и нимф. Внутри агрегации клещи присасываются столь тесно, что налегают друг на друга наподобие черепицы, а в коже, вокруг их ротовых органов, образуется общий воспалительный очаг. Одновременное питание многих особей на ограниченном участке тела, по-видимому, не мешает нормальному насыщению паразитов. Формирующийся в зоне прикрепления обширный очаг геморрагического воспаления обеспечивает кровяное питание иксодид уже с первых часов или дней прикрепления. Напротив, при питании клещей на животных с приобретенной устойчивостью к этим паразитам гнойный характер воспаления вызывает серьезные нарушения в процессе питания. Это явление было описано В. А. Мусатовым (1970) в опытах с кормлением Н. marginatum, Н. asiaticum и R. bursa на овцах и кроликах.
Формирование подобных агрегаций обеспечивается механизмами феромонной коммуникации. Для нескольких видов рода Amblyomma, включая A. hebraeum и A. variegatum, установлено, что самцы после нескольких дней питания начинают выделять летучие феромоны скопления-прикрепления. Они воспринимаются голодными самками, а также самцами и, возможно, неполовозрелыми особями с расстояния до 4 м. Клещи направленно движутся в сторону хозяина, а после прицепления к нему перемещаются по телу к ранее прикрепившемуся самцу, испускающему феромоны. Наряду с направленным движением феромоны стимулируют и сам процесс прикрепления, которое происходит в непосредственной близости от питающегося самца (Norval et al., 1991b, 1991c). У других изученных в этом плане видов из родов Dermacentor, Rhipicephalus, Hyalomma, Aponomma привлекающим объектом служат питающиеся самки, которые выделяют половые аттрактанты. Под их влиянием самцы, питавшиеся 3—5 сут и способные к копуляции, открепляются, находят готовых к спариванию самок, копулируют с ними и затем повторно присасываются вблизи от них (Sonenshine, 1985).