Иксодовые клещи — паразиты и переносчики инфекций
Прием пищи
Началом собственно питания следует считать момент, когда клещ прорезает кожу хозяина и погружает в нее хелицеры и гипостом. До этого попавший на хозяина клещ может перемещаться по его телу от нескольких минут до нескольких часов в поисках места прикрепления. Выбор места прикрепления осуществляется с помощью пальпальных хеморецепторов и механорецепторов I пары ног. Важную роль в этом процессе могут играть и механорецепторы пальцев хелицер, тестирующие поверхность места прикрепления и поверхность кожи в процессе ее прорезания. Кроме механорецепторов на хелицерах нескольких видов из родов Boophilus, Hyalomma, Dermacentor, Haemaphysalis, Ixodes обнаружены хеморецепторы (рис. 17; табл. XXIX, 4), выполняющие функции органов вкуса (Waladde, Rice, 1982; Данилов, 1987а, 19876, 1988). Как и у других кровососов, фагостимуляторами для питающихся клещей служат АТФ и глютатион (Paine et al., 1983). В то же время гистамин и некоторые другие вещества могут прерывать питание и даже вызывать открепление клещей (Binnington, Kemp, 1980; Losel et al., 1992).
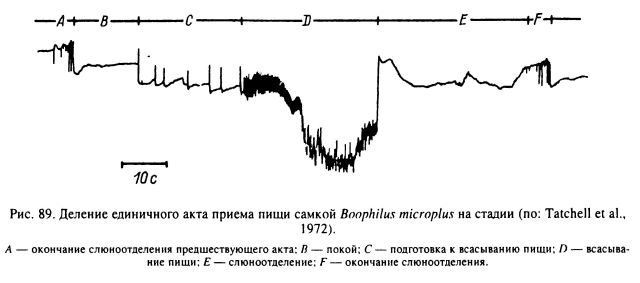
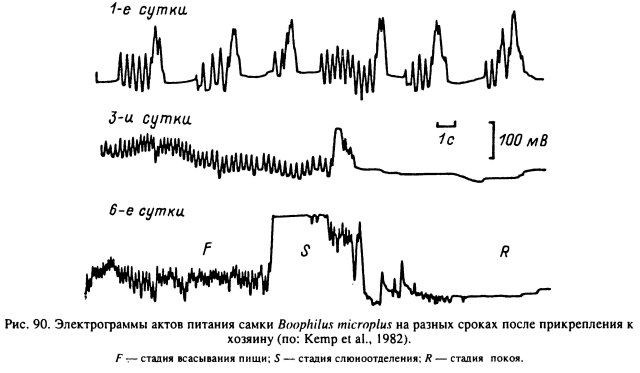
Прямыми наблюдениями над питающимися особями и с помощью электрофизиологических методик для самок D. andersoni (Gregson, 1967, 1969) и В. microplus (Tatchell et al., 1972) было установлено, что процесс многодневного питания складывается из множества элементарных актов всасывания жидкой пищи и впрыскивания слюны, разделенных паузами неактивности. Фаза всасывания слагается из ритмических, от 5 до 15 и более в секунду, расширений-сокращений глотки, проталкивающих жидкое содержимое через пищевод в среднюю кишку. Затем обязательно следует фаза одномоментного выталкивания слюны в предротовую полость, и наступает фаза неактивности с временной приостановкой всасывания и слюноотделения (рис. 89). За время питания продолжительность элементарных актов питания и его составных частей меняется в сторону удлинения, и максимальная активность наблюдается в последние сутки питания, когда у самок В. microplus - фаза неактивности сокращается до 15 % времени питания (рис. 90).
В случае прикрепления клещей к несенсибилизированным животным уже в первые минуты после проникновения ротовых частей в кожу наблюдается местное расширение и повышение проницаемости стенок кровеносных капилляров. Вследствие этих процессов у дистальных концов хелицер могут формироваться небольшие гематомы, из которых клещ получает первые порции пищи. Одновременно развивается воспалительный отек, который распространяется на всю толщу кожи, включая эпидермис, и клещ может быть окружен невысоким кольцевым валиком.
Капилляры первоначально заполняются нейтрофилами и в меньшей степени лимфоцитами. Эти клетки проходят сквозь стенки сосудов и мигрируют в направлении окончания гипостома и хелицер. В зоне инфильтрации нейтрофилов происходит расширение дермы и формируется так называемая пищевая полость (табл. XXXVII), из которой клещ отсасывает ее жидкое содержимое (Tatchell, Moorhouse, 1968). Содержимое пищевой полости меняется в зависимости от фазы питания и особенностей воспалительной реакции хозяина. Размеры пищевой полости в процессе питания увеличиваются и к концу питания A. americanum на морских свинках достигают у личинок 275 х 305 мкм, у нимф — 467 х 567 и у самок — 1900x1717 мкм (Brown, Knapp, 1980а, 1980b). При питании самок R. sanguineus на собаках размеры пищевой полости составляют в среднем 600x1275 (Theis, Budwiser, 1974) и самок R. appendiculatus на крупном рогатом скоте и кроликах — 3500x1000 мкм (Walker, Fletcher, 1986).
На фазе роста значительную часть содержимого пищевой полости и соответственно полости средней кишки могут составлять некровяные компоненты воспалительного инфильтрата, включая нейтрофилы и в меньшем количестве базофилы, эозинофилы и другие мигрировавшие клетки. В конце питания, благодаря увеличению проницаемости и разрушению стенок окружающих капилляров и кровеносных сосудов, пищевая полость заполняется цельной кровью и фактически превращается в обширную гематому. Так, для клеща В. microplus с помощью радиоактивного мечения эритроцитов хозяина было показано, что за первые 6 сут питания самки поглощали в среднем около 3.8 мкл крови, что составляло меньшую часть от общего объема пищи. За последние сутки питания эти клещи выпивали в среднем 300—500 мкл цельной крови (Seifert et al., 1968).
Характер кожных реакций хозяев может меняться в зависимости от их видовой принадлежности, но часто клещи успешно питаются на, необычных для них хозяевах. Характер кожных реакций в этих случаях может существенно не отличаться, как например при питании самок R. appendiculatus на крупном рогатом скоте и морских свинках (Walker, Fletcher, 1986), Некоторые виды хозяев не подходят для питания клещей в связи с интенсивной пролиферацией эпидермиса, как описано для личинок и нимф Hyalomma при паразитировании на земляных крысах (Nesokia indica). Из-за быстрого размножения эпидермальных клеток клещи оказываются замурованными в кожу грызуна и погибают (Лотоцкий и др., 1959). Иногда препятствием для нормального насыщения клещей могут служить мелкие размеры хозяина. Например, при питании самок I. persulcatus на мелких грызунах острый воспалительный процесс может возникнуть уже в начале питания и препятствовать насыщению клещей (Калягин, 1976).