Иксодовые клещи — паразиты и переносчики инфекций
Взаимоотношения с хозяевами
Рассмотренный выше сложный комплекс взаимодействий питающихся клещей с их хозяевами определяет многие важные особенности образуемой ими паразитарной системы. Именно на паразитических стадиях развития обеспечивается регуляция численности иксодид в зависимости от их плотности, так что в природных экосистемах преобладает относительно невысокая численность клещей на их главных хозяевах при умереннойх напряженности противоклещевого иммунитета. В результате паразитирование клещей в большинстве случаев не ведет к гибели хозяев, а иммунные реакции не исключают возможности повторных питаний на сенсибилизированных животных (Балашов, 1992а).
Количественным оценкам взаимодействия клещей с хозяевами препятствуют существенные пробелы в установлении их вредоносности. Пока мы можем оперировать лишь случаями высоких уровней заклещевленности, когда наблюдаются гибель хозяев или заметные нарушения в их жизнедеятельности. В литературе описаны случаи падежа диких животных вследствие массового паразитирования на них клещей, как например антилоп в национальном парке Зимбабве, пораженных R. appendiculatus (Norval, Lightfoot, 1982), лосей в Канаде, когда на одном животном было свыше 100 000 нимф и имаго D. albipictus (Samuel, Barker, 1979), и молодняка белохвостых оленей в США от нападений A. americanum (Bloemer et al.f 1988).
Предельные количества клещей, которые одновременно могут прокармливаться на одном животном, зависят не только от соотношения размеров тела хозяина и паразита, но и от их видовых особенностей. По наблюдениям В. А. Мусатова (1970), при кормлении половозрелых R. bursa на овцах тяжелая интоксикация и гибель животных наблюдались при одновременном питании 3—4 самок на 1 кг массы тела, а в случае кроликов даже при питании 10 клещей на 1 кг хозяева оставались живыми. В случае с нимфами D. reticulatus (Олсуфьев, Каграманов, 1947) обыкновенные полевки (масса тела 25 г) гибли от одновременного питания на них 25—50 клещей, а рыжие полевки с близкой массой тела — от 16 клещей. У отловленного в природе взрослого самца обыкновенной полевки с 312 присосавшимися нимфами были обнаружены многочисленные клинические признаки серьезного заболевания и истощения (Олсуфьев, 1953).
Наибольшая вредоносность иксодовых клещей определяется общим истощением позвоночных животных от потерь крови и особенно от интоксикации слюной. В ветеринарной литературе описаны многочисленные случаи угнетения роста и гибели овец при паразитировании на них нескольких сотен имаго D. marginatus, R. bursa и других видов (Марков, Курчатов, 1949; Мусатов, 1970). В эксперименте наряду с потерями массы тела развивалась анемия у крупного рогатого скота от питания на нем В. microplus (Springell et al., 1971), у овец и кроликов — от Н. marginatum и R. bursa (Мусатов, 1970), кроликов и рыжих полевок — от I. ricinus (Лабецкая, 1990) и лосей — от D. albipictus (Glines, Samuel, 1989). У животных были отмечены значительное уменьшение количества эритроцитов и гемоглобина в крови, лейкоцитоз, гипоальбуминия и другие изменения биохимического состава сыворотки.
Учитывая крупные размеры тела иксодид, особенно их самок, а также поглощение ими во время питания массы крови, в несколько раз превышающей таковую тела напитавшейся особи, прямые потери от их паразитирования могут быть весьма значительными. Самки наиболее крупных видов из родов Amblyomma, Hyalomma и Dermacentor в среднем поглощают за время питания до 8—10 мл крови и более, мелкие виды, как например В. microplus, H. longicornis или R. bursa, — 0.7—1.5 мл (см. главу VI).
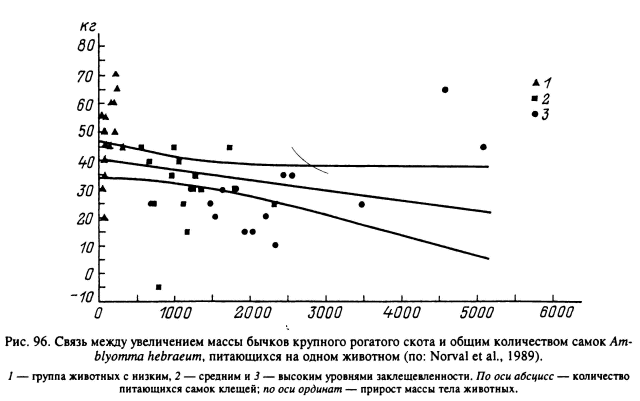
На одной особи крупного рогатого скота может одновременно паразитировать несколько десятков самок, а всего в течение года — несколько сотен клещей. Легко подсчитать, что суммарные потери крови могут измеряться в подобных случаях литрами. Для сельскохозяйственных животных приводятся факты снижения привесов и истощения, но только при паразитировании на них достаточно большого числа клещей (рис. 96). Так, в Южной Африке отмечены заметные потери массы крупного рогатого скота при одновременном паразитировании на нем в течение нескольких месяцев не менее 50 имаго (Young et al., 1988). В серии специальных экспериментов подсчитано, что для крупного рогатого скота завершение питания на животном одной самки A. hebraeum эквивалентно потере 10±4 г крови (Norval et al., 1989), R. appendiculatus — 4 г (Norval et al., 1988b), B. microplus — 0.6—1.5 г (Sutherst et al., 1983). Паразитирование клещей не всегда ведет к заметным потерям массы. Например, питание на скоте личинок, нимф и самцов A. hebraeum существенно не влияло на привесы животных (Norval et al., 1989).
Некоторые из биологически активных компонентов слюны являются нетоксичными для позвоночных животных. Наиболее детально исследованы токсины слюны, вызывающие клещевые параличи, которые к настоящему времени выявлены при питании 30 видов иксодид на различных животных и человеке (Gothe et al., 1979). Наибольшую опасность представляют D. andersoni и D. variabilis в Северной Америке, I. rubicundis в Южной Африке и I. holocyclus в Австралии. Параличи развиваются только при паразитировании не менее определенного количества клещей, разного для разных видов хозяев, их возраста и массы тела (Fourie et al., 1989). Они могут исчезнуть при своевременном удалении всех питающихся паразитов. У клещей I. holocyclus из слюнных желез удалось изолировать паралитический токсин белковой природы с молекулярной массой 40 000—60 000 (Stone, Binnington, 1989). Клещевым токсикозом, связанным со слюной, является также широко распространенная в Южной Африке «болезнь потливости» крупного и мелкого рогатого скота, свиней и собак, часто возникающая при питании клеща Н. truncatum (Spickett, 1992).
Существенный вред хозяевам могут причинить разрушения тканей в местах прикрепления клещей. При питании одиночных клещей на нативных или на умеренно сенсибилизированных животных в местах прикрепления паразитов преобладают геморрагичесие синдромы с умеренной клеточной инфильтрацией и ограниченными пролиферативными процессами. Заживление ранки по отпадении клеща протекает без осложнений с образованием струпа и замещением пищевой полости гранулированной тканью (Мусатов, 1978). Реже питание сопровождается гнойно-некротическим
воспалением, иногда осложняющимся вторичной стафилококковой инфекцией (Webster, Mitchell, 1989). Места прикрепления клещей могут служить очагами для откладки яиц и развития личинок миазных мух. Чаще гнойно-некротические поражения случаются при одновременном нахождении на ограниченном участке тела многих клещей, как в случае питания Н. marginatum или R. bursa на спине овец, у которых в дерме развивались обширные полости, заполненные почти одной кровью, гнойно-некротической массой или сгустками агглютинированных эритроцитов. В конечном итоге верхние слои кожи отторгались и оставались глубокие, долго незаживающие раны (Мусатов, 1978). У молодняка белохвостого оленя в США питание 150—300 имаго A. americanum на голове, ушах и вокруг глаз вело к некрозу окружающих тканей, ослеплению и гибели животных (Bloemer et al., 1988). Остающиеся дефектными места питания клещей, особенно их крупных видов, являются одной из главных причин снижения сортности кожевенного сырья и значительных экономических потерь (Кербабаев, Скосырских, 1988).
Наибольший вред от паразитирования клещей испытывают сельскохозяйственные животные, отличающиеся высокой плотностью на единицу площади пастбища. Сходная ситуация иногда возникает и в разного рода заповедниках, где также перестают действовать естественные механизмы регуляции численности паразитов. В ненарушенных экосистемах численность иксодовых клещей на диких животных-хозяевах обычно умеренная или низкая, а ущерб от их паразитирования невелик.
Численность клещей на хозяевах чаще всего оценивают по двум показателям — индексам обилия и встречаемости, которые соответственно отражают среднюю пораженность хозяев этими паразитами и процент пораженных особей в популяции. Детальные исследования численности клещей на их хозяевах в природных местообитаниях выполнены для многих видов, и к их числу принадлежит I. persulcatus. На мелких млекопитающих и птицах обычно одновременно паразитирует не более 3—4 личинок и нимф этого вида, а на средних по размерам тела млекопитающих (зайцах, ежах, хищниках) может быть более 10. Главными прокормителями имаго служат дикие животные и скот. На этих животных индекс обилия обычно не превышает 10 самок и лишь в некоторых регионах может достигать 100 особей (Таежный клещ..., 1985).
Заклещевленность отдельных особей хозяев может быть значительно выше средних значений. Например, на лесном коньке или овсянке-ремезе (масса тела каждого около 20 г) и на дроздах (масса тела 60—80 г) одновременно паразитировало до 70 личинок и нимф, а на рябчике (масса тела 400 г) — до 130 неполовозрелых особей. На отдельных экземплярах землероек находили до 47 личинок и нимф, на мышевидных грызунах — до 363 личинок и 44 нимф, а на белках — до нескольких сотен. На еже или зайце, по данным разных авторов, может одновременно паразитировать до 200—300 личинок и нимф, а также до 40 и более самок (Таежный клещ..., 1985). На коровах максимальное количество одновременно питающихся самок достигало на Алтае 534 на одно животное (Дмитриев, 1962).
Сходны с таежным клещом трофические связи и у другого широко распространенного вида — I. ricinus. Местами его численность очень высокая и пораженность прокормителей может достигать 80—100 %. В разных регионах индекс обилия имаго на крупном рогатом скоте значительно варьирует. Например, в Молдавии этот показатель колеблется от 3.03 до 39.77 самок (табл. 26) (Успенская, 1987), в Новгородской области — от 0.1 до 118 (Померанцев, 1935), в южной части Ленинградской области — от 0.24 до 50.7 (Алфеев, 1935). Максимальное число самок, одновременно паразитировавших на одной корове, достигало 188 (Померанцев, 1935) и даже 256 особей (Арзамасов, 1961).
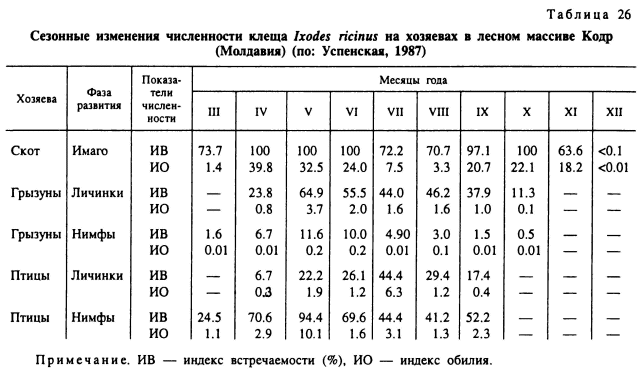
Весьма значительно варьируют и индексы обилия неполовозрелых фаз. Так, в разных частях Молдавии на мышевидных грызунах усредненные годовые индексы обилия личинок I. ricinus были 0.21—7.67 при индексах встречаемости 14.0— 97.43 %, а нимф — соответственно 0.02—1.70 и 2.08—58.3 % (Успенская, 1987). В Белоруссии, при более низкой численности этого вида средний индекс обилия личинок и нимф на грызунах составлял 1.3 при встречаемости 29.7 %. Там же встречаемость личинок и нимф этого вида на ежах составила 70.6 % и индекс обилия — 41.3, на кротах — 11.3 % и 4.9 и на обыкновенной бурозубке — 19.3 % и 3.6 соответственно. Обращает на себя внимание очень сильная заклещевленность ежей, среди которых отмечена одна особь, пораженная 389 личинками и нимфами. На одном кроте может паразитировать до 40 клещей и на обыкновенной бурозубке — до 38. На одной особи рыжей полевки зарегистрировано паразитирование 52 личинок, на желтогорлой мыши — 50, на белке — 132 личинки или 199 нимф (Арзамасов, 1961). В одном из очагов энцефалита в Австрии (Pretzmann et al., 1964) в среднем за сезон на одной желтогорлой мыши прокармливается 19.9—27.9 личинок и 0.17—0.56 нимф, на лесной мыши — 19.9—27.9 и 0.17—0.56, на рыжей полевке — 14.8—20.8 и 0.36—0.41 соответственно.
Данные о заклещевлении диких копытных этим видом немногочисленны из-за сложности отстрелов хозяев в сезон активности клещей. Согласно Б. П. Савицкому (1985), в Беловежской Пуще (Белоруссия) индексы обилия самок на лосях 7.31, на косулях — 4.64 и на кабанах — 2.53.
При отсутствии противоклещевых мероприятий весьма высокой может быть численность клеща D. reticulatus. По многолетним наблюдениям Н. Г. Олсуфьева (1953) на юге Московской области, при средней численности голодных имаго от 1120 до 2450 особей/га в период весеннего пика активности на крупном рогатом скоте, в разных стадах и в разные годы, в среднем на одном животном одновременно паразитировало от 0.3 до 189 самцов и самок. Максимальное же количество питавшихся клещей составляло 654 особи.
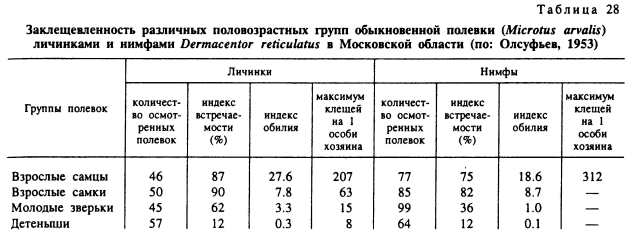
Весьма высокой в этом районе была и численность неполовозрелых фаз на мелких млекопитающих. Из 17 видов-прокормителей личинок и нимф главными хозяевами служили обыкновенные полевки (Microtus arvalis) и обыкновенные бурозубки (Sorex araneus), на которых прокармливалось свыше 80 % всех клещей при соответственно 67—85 и 77—93 % пораженных зверьков в период пика численности паразитов. Усредненные индексы обилия личинок в разные годы варьировали для полевок в пределах 3.8—9.6 особей и нимф — 1.5—5.7. Максимальная заклещевленность составила 652 личинки или 312 нимф на 1 полевку. У бурозубок индексы обилия личинок были 4.8—12.8 и нимф — 2.2—4.7 при максимальных значениях в 181 личинку и 41 нимфу.
У широко распространенного в США D. variabilis, по данным многолетних наблюдений в Виргинии (Sonenshine, 1975), в условиях питания подавляющей массы клещей на диких животных индексы встречаемости и обилия были значительно ниже, чем у D. reticulatus в России. 96 % личинок и 93.5 % нимф этого вида прокармливалось на белоногом хомячке (Peromyscus leucopus) и пенсильванской полевке (Microtus pennsylvanicus). Усредненные индексы обилия за 7 лет наблюдений для хомячков составили 2.3 личинки и 0.15 нимфы, а для полевок соответственно 4.4 и 0.64. Суммарные индексы обилия на грызунах для обеих фаз варьировали в разные годы от 2.30 до 8.30 клещей. Половозрелые клещи питались преимущественно на млекопитающих со средними размерами тела. Около 50 % из них прокармливалось на енотах-полоскунах, и за 5-месячный период активности имаго индекс обилия их на одном еноте в среднем составил 31.9 самцов и самок. Для опоссумов (Didlephis marsupialis) индекс обилия составил 9.78 клещей, для скунсов (Mephitis mephitis) — 8.72 и для красных лисиц (Vulpes fulva) — 10.23.
В степях, пустынях и саваннах обитающие там представители родов Rhipicephaius, Hyalomma, Amblyomma и Boophilus на имагинальной фазе, а у некоторых видов также на личиночной и нимфальной, паразитируют на диких копытных, но преимущественно на крупном и мелком рогатом скоте, лошадях и верблюдах. Несмотря на то что на этих хозяевах одновременно или последовательно в разные сезоны года может питаться несколько видов клещей, средние суммарные индексы обилия этих паразитов редко превышают 100 имаго на одну особь хозяина.
По многолетним данным индексы обилия для 14 видов иксодид, встречающихся на скоте в Туркмении, составили для крупного рогатого скота 22.4 особи при индексе встречаемости 52.5 %, для овец — 4.9 и 58.6 %, для коз — 8.35 и 79.8 %, для верблюдов — 22.5 и 79.1 % соответственно (Бердыев, 1980). В Судане при совместном паразитировании на крупном рогатом скоте местной породы клещей A. lepidum, A. variegatum, В. decoloratus, H. anatolicum, R. sanguineus на различных стадиях индексы обилия варьировали от 17.6 до 40.5 самцов и самок, и только среди животных завозного происхождения (Jongejan et al., 1989) этот показатель достигал 71.5. В Замбии на крупном рогатом скоте паразитирует 12 видов клещей, и их индексы обилия в разных стадах варьировали от 1.9 до 85.6 особей (McLeod et al., 1977). В Юго-Западной Африке при преимущественном паразитировании на скоте Н. marginatum rufipes и Н. truncatum индексы обилия в разных стадах варьировали от 6.9 до 286.6 клещей (Biggs, Langenhoven, 1984).
В Танзании при доминировании на скоте R. appendiculatus в период пиков активности имаго индексы обилия составляли в среднем 100—120 самцов и самок (Tatchell, Easton, 1986), а в Уганде численность клещей выше — до 500 (Kaiser et al., 1982). В Австралии, где основная масса клещей на скоте представлена В. microplus, в стадах из животных с разной долей крови азиатского и зебувидного скота индексы обилия самок варьировали от 37—79 до 302—465 клещей (Bourne et al., 1988).
На отдельных особях прокормителей количество одновременно питающихся клещей может быть значительно выше средних величин для стада или популяции. Например, при среднегодовом индексе обилия 9.55 наибольшее число самцов и самок Н. asiaticum на одной корове достигало 318, а нимф — более 1832 особей при индексе обилия 142.5 (Бердыев, 1980). Максимальное число нимф и имаго Н. scupense на одно животное составило 4682 особи (Марков, Курчатов, 1949). В Кении при одновременном паразитировании на скоте имаго нескольких видов иксодид максимальная заклещевленность достигала 750 особей (Tatchell, 1986), в Танзании — 901 (Tatchell, Easton, 1986), в Замбии — 1072 (Jongejan et al., 1989). В Монголии с одной коровы было снято 3116 имаго D. nuttali, а с лошади — 1725 (Dash, 1988).
Данные о максимальной численности иксодовых клещей на диких животных в значительной степени фрагментарны. Известно, что в Северной Америке на одном взрослом лосе могут одновременно питаться свыше 100 000 личинок, нимф и имаго однохозяинного D. albipictus, и это служило причиной гибели части пораженных животных (Samuel, Barker, 1979). На антилопе-бушбуке (Tragolaphus reriptus) количество одновременно питающихся клещей H. silacea составляло 515 самцов, 21 самку, 345 нимф и 3645 личинок (Norval, 1975a).
Весьма большой может быть численность клещей на млекопитающих со средними размерами тела. С одного барсука в Туркмении было снято 464 имаго R. turanicus (Бердыев, 1980), а в Молдавии более 500 личинок, нимф и имаго I. crenulatus (Успенская, 1987). В Австралии на главном прокормителе клеща I. holocyclus — бандикуте (Isoodon macrourus) нормально заканчивают питание от 500 до 2000 личинок, 100—200 нимф и 4—6 самок (Doube, 1979).
На средних по размерам тела пресмыкающихся также может одновременно паразитировать большое количество клещей. По нашим наблюдениям, на степных черепахах (Testudo horsfieldi) в предгорных районах Таджикистана питались до 9—12 самок и 20—25 самцов или 80—90 нимф Н. aegyptium. В Туркмении в горах Кугитангтау максимальное число Н. sulcatd на горных агамах достигало 37 нимф.
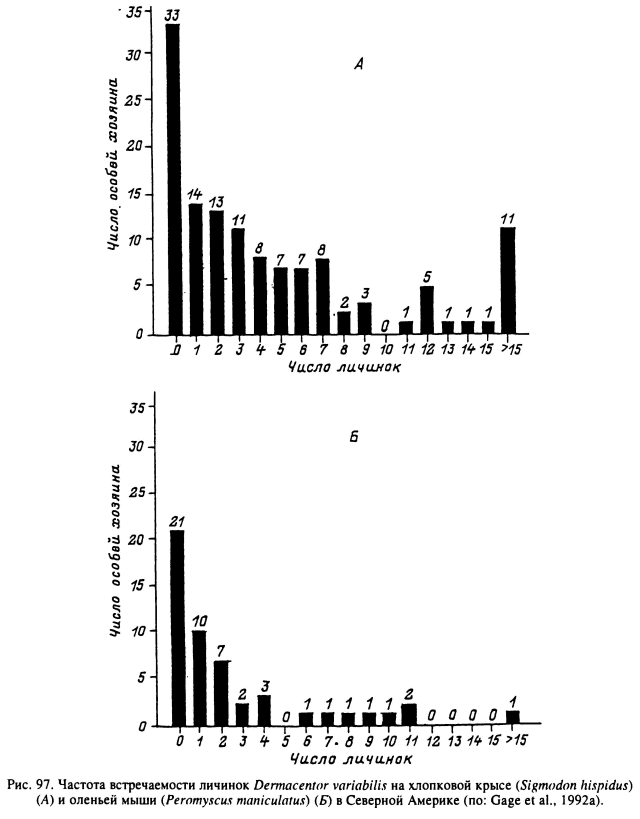
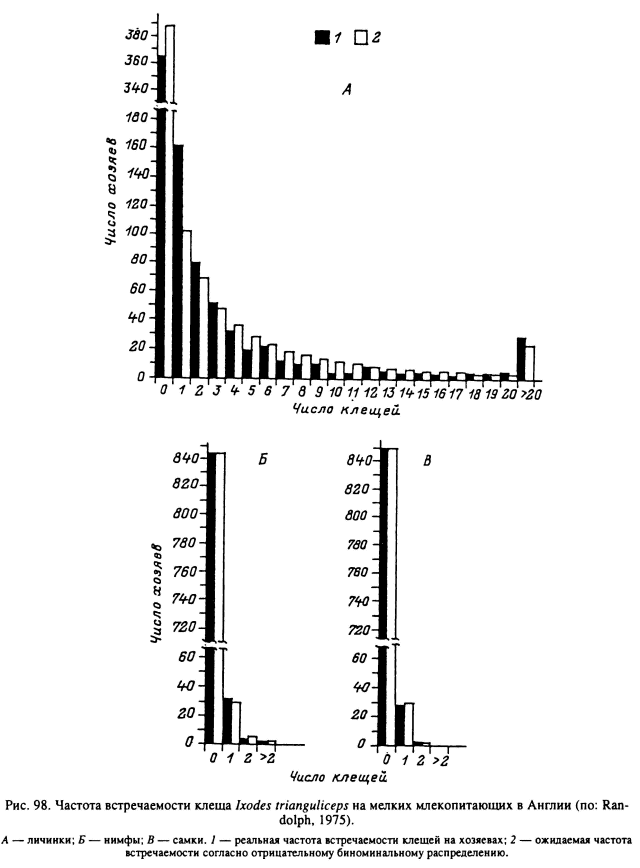
При изучении особенностей распределения иксодовых клещей внутри популяций их хозяев было выяснено, что наиболее часто приводимые индексы обилия и встречаемости оказываются недостаточно нормативными. В немногих работах, специально посвященных этой проблеме, было обнаружено, что меньшая часть популяции хозяина в большинстве случаев прокармливает большую часть популяции клещей. Подобное распределение свойственно для многих групп паразитов, и иксодовые клещи не составляют исключения. Впервые на это обратил внимание К. А. Бреев (1972), обнаруживший, что распределение питающихся личинок клещей I. ricinus на лесной мыши агрегированного типа близко к отрицательному биноминальному. Подобный же тип распределения личинок этого вида был обнаружен и на мелких млекопитающих в Швеции (Nilsson, Lundqvist, 1978), у личинок и нимф D. variabilis в США (рис. 97) (Gage et al., 1992) и у всех фаз развития I. trianguliceps в Англии (рис. 98) (Randolph, 1975). По наблюдениям Олсуфьева (1953), 70 обыкновенных полевок из 262 осмотренных особей прокармливали на себе 46 % от общего числа (6930) собранных с них личинок D. reticulatus. При регулярных обследованиях на заклещевленность одного из стад крупного рогатого скота в Уганде (Kaiser et al., 1982) на 25 % животных прокармливалось свыше 50 % самок A. variegatum. На особях, входящих в состав указанных 25 %, прокармливалась также большая часть самцов, нимф и личинок этого же вида, а также других сопутствующих иксодид. Подобное же агрегированное распределение имаго R. evertsi обнаружено на группе из 14 коров, выпасавшихся совместно на одном пастбище в Танзании (Tatchell, 1986). На одном из этих животных клещи полностью отсутствовали, на трех паразитировало в среднем 1—5 клещей, на пяти — 6—10, на трех — 11 —15 и еще на двух коровах — 16—20 и 21—25 клещей.
Отрицательное биноминальное распределение было свойственно клещам A. hydrosauri и A. limbatum, все стадии которых могут одновременно паразитировать на ящерице Trachydosaurus rugosis в Австралии (Bull et al., 1989). Из 502 осмотренных ящериц более 50 % были свободны от паразитов, а большая часть клещей прокармливалась на 10—20 % рептилий. В это число входило 5 ящериц, на которых одновременно питалось более 100 личинок и 40 нимф A. hydrosauri, a также 3 ящерицы, прокармливающие свыше 100 личинок и 29 нимф A. limbatum. Сходное агрегированное распределение обнаружено нами и у нимф Н. sulcata на горных агамах в горах Кугитангтау (Туркмения). Из 114 осмотренных в конце апреля ящериц клещи были обнаружены на 32. Из собранных с них 390 клещей 310 питались на 10 ящерицах, что по отношению ко всем осмотренным особям составило около 3.2 %. При паразитировании на других видах рептилий агрегированное распределение выявлено у клещей I. ricinus (Bauwens et al., 1983), I. asanumai (Hayashi, Hasegawa, 1984), I. uriae (Boulinier et al., 1996) и других видов. Неслучайный агрегированный тип распределения иксодид на хозяевах может быть обусловлен многими причинами, среди которых следует различать внешние по отношению к организму хозяина и присущие последнему. К первым следует отнести агрегированное размещение голодных личинок, образующих скопления в непосредственной близости вокруг кладки яиц. Подобное агрегированное размещение голодных личинок на растительности было описано для I. persulcatus в таежных лесах (Левин, 1987), I. uriae — в колониях морских птиц (Boulinier et al., 1996) и В. microplus — на открытых пастбищах Австралии (Zimmerman, Garris, 1987). Однако для нимф и имаго, несмотря на возможность образования скоплений в местах преимущественного отпадения напитавшихся особей предшествующей фазы на лежках хозяев вдоль их троп и в других, более часто посещаемых биотопах, размещение голодных клещей ближе к случайному. Поэтому встречаемость и обилие клещей на хозяевах не могут быть объяснены только топографией размещения голодных стадий этих паразитов на местности.
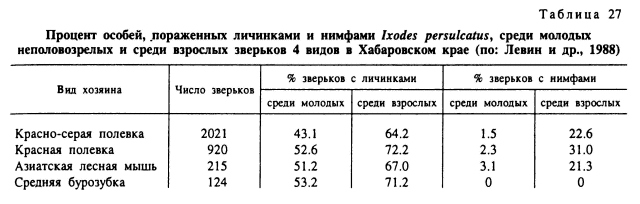
Первостепенное значение в неравномерном заклещевлении особей одной популяции хозяина играют особенности поведения отдельных индивидов, включая разную двигательную активность, иерархический статус в стаде, привлекательность для клещей и др. Эти особенности хозяина зависят от возраста, пола и индивидуальных различий в типе нервной системы. В частности, многими авторами отмечается большая заклещевленность самцов мелких млекопитающих по сравнению с молодняком и самками (табл. 27, 28). Среди крупного рогатого скота часто наблюдается постоянное преимущественное питание клещей на одних и тех же животных в стаде (Олсуфьев, 1953; Sonenshine, 1975; Tatchell, 1986; Окулова, 1986). Эти и многие другие факторы определяют шансы встречи клеща с хозяином (рис. 99). После того как клещ оказался на его теле, главенствующая роль в выживании и успешном завершении питания принадлежит уже факторам организма хозяина, его морфологическим, биохимическим особенностям, но в первую очередь его иммунному статусу. Вследствие значительного генетического полиморфизма природных популяций позвоночных животных существуют большие индивидуальные различия по многим морфологическим и биохимическим параметрам, и именно они определяют разную выживаемость клещей на особях одной популяции или стада сельскохозяйственных животных.
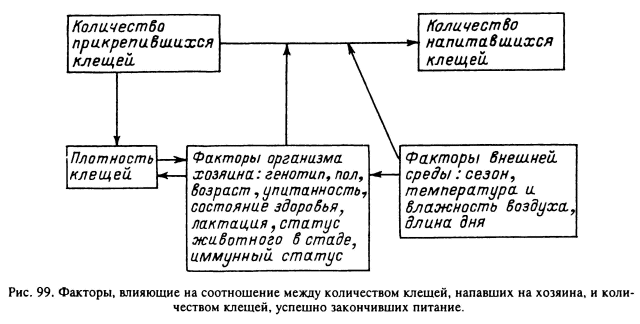
Отношение числа успешно напитавшихся клещей к общему числу особей, попавших на тело хозяина, представляет итог взаимодействия многих факторов, среди которых первостепенное значение имеют «противоклещевые» реакции хозяина от простого механического самоочищения до комплекса иммунных реакций. Паразитарная система иксодовый клещ—хозяин обычно относительно стабильна в пространстве и времени. Численность клещей, одномоментно паразитирующих на их главных прокормителях, в большинстве случаев значительно ниже уровней достоверной вредоносности и тем более — летальности. При этом годовые колебания индексов обилия клещей значительно меньше, чем колебания численности их хозяев. По-видимому, существуют два главных механизма регуляции численности клещей. Один из них связан с многообразными воздействиями окружающей среды, и в первую очередь температуры и влажности, на свободноживущие стадии. Известны многочисленные случаи гибели голодных особей при засухе, а напитавшихся, кроме того, и при затоплении. Однако в подобных случаях доля погибших особей не связана с общей численностью популяции клеща и прокормителя.
Стабильность паразитарной системы, по нашему мнению, обеспечивается сложным комплексом паразито-хозяинных взаимодействий на стадии питания. Одним из результатов подобных взаимодействий является сенсибилизация позвоночного животного антигенами слюны клеща и развитие состояния резистентности. В естественных условиях главные хозяева, несмотря на постоянный контакт с клещами, не обладают абсолютной устойчивостью, так что часть клещей может успешно завершить питание и на сенсибилизированных особях. С другой стороны, значительная часть популяции хозяина может быть слабо поражена этими паразитами, и благодаря агрегированному распределению большая часть клещей может прокармливаться на немногих особях хозяина.
Агрегированный тип распределения питающихся иксодовых клещей внутри популяции хозяев, по-видимому, служит одним из механизмов регуляции их численности, в зависимости от плотности на хозяевах, действующим по принципу обратной связи. При высокой численности паразита немногие особи популяции хозяина, особенно подверженные заклещевлению, не могут выдержать клещевую инвазию и погибают. Однако вместе с ними гибнут и не закончившие питание клещи, могущие составлять значительную долю всей популяции. В результате стабильность паразитарной системы может поддерживаться в сбалансированном состоянии при умеренной плате хозяев за «паразитарный груз».
Подобные равновесные состояния свойственны естественным экосистемам. В экосистемах, подвергнутых антропогенной трансформации или экологическому стрессу, механизмы взаимной регуляции численности партнеров по паразитарной системе нарушаются. Некоторые виды иксодид, приспособившиеся к новым условиям существования и питанию на скоте или на немногих видах диких животных, могут достигать необычных для природы уровней численности и вызывать падеж или причинять существенный ущерб здоровью своих новых прокормителей. Неблагоприятная ситуация часто усугубляется слабыми иммунными реакциями у завозных животных против местных видов паразитов.