Иксодовые клещи — паразиты и переносчики инфекций
Глава VIII
Самкам иксодовых клещей свойственна жесткая гонотрофическая гармония, так как необратимые морфофизиологические изменения в их организме во время питания и подготовки к яйцекладке делают невозможным прохождение повторных гонотрофических циклов. Нормальное завершение гонотрофического цикла связано с успешным насыщением, которое возможно только у осемененных самок.
Встреча полов. Самки I. ricinus, I. persulcatus (Балашов, 1967; Graf, 1978b), I. scapularis (Yuval, Spielman, 1990), I. rubicundus (Fourie, Kok, 1995), I. trianguliceps (Randolph, 1980) и, вероятно, других видов рода Ixodes способны к копуляции в голодном состоянии. Это неоднократно наблюдали как в природе, так и в лабораторных культурах клещей. В природных популяциях I. persulcatus среди самок, собранных с растительности, по данным вскрытий, осемененные особи составляли от 0 до 50—65 %. При этом за сезон активности клещей доля осемененных самок постепенно увеличивается. Этот же показатель возрастал при увеличении плотности клещей и особенно обилия активных особей (Успенский, Репкина, 1978; Бабенко и др., 1979). У I. ricinus доля осемененных самок, собранных с растительности в Швейцарии, варьировала от 27 до 100 % (Graf, 1978b) и в Латвии — 22—30 % (Бабенко и др., 1979). Высокий процент осемененных самок выявлен также среди активных голодных особей I. rubicundus и I. scapularis. На хозяев нападают осемененные и девственные самки видов этого рода, а также самцы. Поэтому осеменение девственных самок возможно и на теле хозяина. В местах присасывания клещей в большом количестве встречаются самцы, копулирующие с питающимися самками.
У видов подсем. Amblyomminae голодные самцы и самки не способны к копуляции. Исключение, возможно, составляют немногие виды Аропотта и Amblyomma, у которых зрелые сперматиды обнаружены в половых путях не питавшихся самцов (Oliver, 1989). У остальных видов голодные самцы не способны к спариванию. Зрелые сперматиды появляются у них на 3—5-е сутки с начала питания. После этого они приступают к копуляции и осеменяют питающихся самок. Голодные самки Amblyomminae также лишены способности к спариванию и осеменению.
У I. ricinus максимальное число копуляций у одного самца было 4, но чаще их бывает только 2. Затем подобные самцы быстро погибают. В то же время девственные самцы живут в благоприятных условиях до года и более (Graf, 1978b). Самцы Amblyomminae способны к значительно большему числу повторных спариваний. В промежутках между спариваниями они повторно прикрепляются к хозяину, питаются и у них возобновляются процессы сперматогенеза. Благодаря подобному восполнению половых продуктов один самец может последовательно осеменить несколько десятков самок. В литературе описано осеменение самцом Н. marginatum 25 самок (Первомайский, 1954), самцами A. americanum — до 27— 37 самок за 68—127 сут (Gladney, Drummond, 1970) и до 22 самок D. variabilis (Oliver, 1982).
Спаривание. Наиболее полно процесс спаривания описан у I. ricinus (Graf, 1978b). После установления самцом контакта с самкой он закрепляется на ее вентральной стороне при помощи II и III пар ног, обеспечивая тесный контакт между вентральными поверхностями партнеров. Ноги самки не участвуют в копуляции, и спаривающиеся самки свободно перемещаются. После закрепления на теле самки самец подводит свои ротовые органы к генитальному отверстию и погружает в него гипостом и хелицеры. Хоботок клеща при этом располагается перпендикулярно к спинному щитку, а пальпы остаются снаружи и лишь раздвигаются плоско в стороны от полового отверстия. В этом положении самец может оставаться несколько часов или даже суток (рис. 100, А).
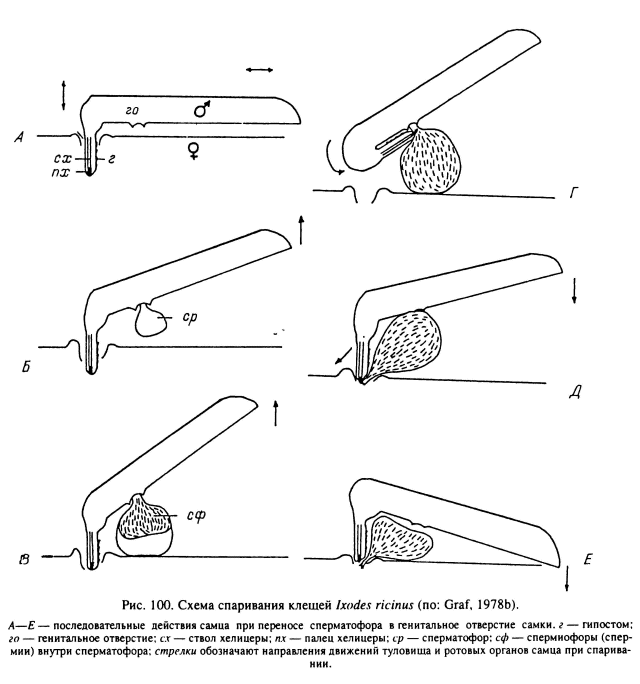
Второй этап копуляции начинается с повторяющихся расширяющих движений хоботка в половом отверстии самки, сопровождающихся выделением капелек слюны. Затем из мужского полового отверстия выходит наружу эктосперматофор, размеры которого быстро увеличиваются за счет заполнения половыми продуктами (рис. 100, Б, В). После этого хоботок выдвигается из полового отверстия самки и подгибается на вентральную сторону, так что хелицеры захватывают шейку сперматофора и переносят его снаружи к женскому половому отверстию (рис. 100, Г, Д). При переносе сперматофора на дистальных концах хелицер видны капельки жидкости, вероятно являющейся секретом слюнных желез. С помощью пальцев хелицер шейка сперматофора вводится внутрь полового отверстия (рис. 100, Е). По завершении копуляции самец оставляет самку.
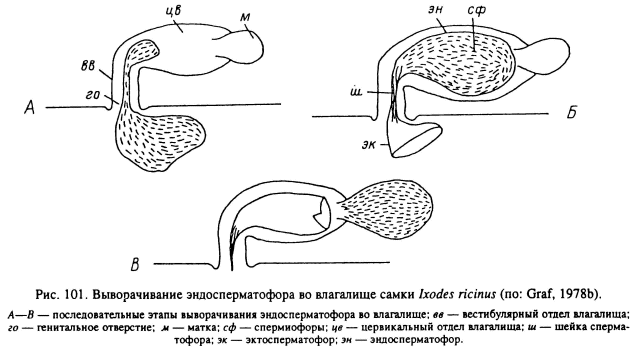
Содержимое эктосперматофора переливается в эндосперматофор, находящийся уже внутри центральной части влагалища. Спустя 48—72 ч после конца копуляции капсула эндосперматофора открывается в своей апикальной области и спермиофоры заполняют матку, размеры которой при этом сильно увеличиваются (рис. 101). Пустой эктосперматофор может оставаться прикрепленным к женскому половому отверстию в течение нескольких часов или дней по окончании спаривания.
У Amblyomminae процесс спаривания в основных чертах сходен с описанным выше для Ixodes, но имеются и определенные различия. Перед спариванием питающийся самец временно открепляется от хозяина в ответ на половые аттрактанты находящейся вблизи самки. В процессе спаривания удержание самцов обеспечивается взаимным зацеплением ног обоих партнеров. В половое отверстие вводятся только пальцы хелицер, а гипостом остается снаружи вместе с пальпами. У некоторых видов (Н. excavatum, R. sanguineus) перенос сперматофора осуществляется скользящими движениями всей идиосомы самца и хелицеры на это время не выводятся из полового отверстия самки. У других видов (A. variegatum, H. marginatum) перенос сперматофора, как и у Ixodes, осуществляется только с помощью ротовых частей (Feldman-Muhsam, Borut, 1983a).
Феромоны. Встреча полов и спаривание у иксодовых клещей регулируются особыми химическими веществами — феромонами. Они выделяются во время питания особями одного или обоих полов, стимулируют соответствующие рецепторы и вызывают групповое прикрепление самцов и самок одного вида на определенных участках тела хозяина.
У видов родов Dermacentor, Hyalomma и Rhipicephalus питающиеся самки выцеляют летучие половые феромоны, главным компонентом которых у всех видов является 2,6-дихлорфенол (Sonenshine, 1985). Местом выделения этого типа феромонов служат особые железы, располагающиеся на аллоскутуме под фовеа. Они начинают функционировать только через несколько суток питания, и их активность совпадает с повышением содержания экдизона в организме. У голодных самок выделение полового феромона отсутствует. Нет реакции на них и у голодных самцов.
Питающиеся самцы воспринимают 2,6-дихлорфенол хеморецепторными сенсиллами передней ямки органа Галлера (табл. XXVI, 3). В ответ на их раздражение питающиеся самцы открепляются и двигаются в направлении питающихся самок. После установления тактильного контакта с последними самцы с готовыми половыми продуктами приступают к спариванию и осеменению самок. Самцы, не достигшие способности к осеменению, повторно прикрепляются к хозяину рядом с питающейся самкой и приступают к копуляции спустя несколько часов или суток по завершении созревания половых продуктов.
2,6-дихлорфенол не обладает видовой специфичностью, и собственно копуляция стимулируется другой группой уже видоспецифичных феромонов. Важная роль в их восприятии принадлежит группе хеморецепторных сенсилл у основания коготка I пары ног (Sonenshine, 1991). В процессе спаривания самцы контактируют пальцами хелицер с половым отверстием самки и окружающей его кутикулой. На этой стадии хеморецепторы пальцев хелицер воспринимают еще одну группу половых феромонов. Последние вырабатываются передними отделами женской половой системы, мало летучи и находятся на поверхности кутикулы. Одним из их главных компонентов являются насыщенные жирные кислоты с цепью из 14—22 атомов углерода. Феромоны этой группы высоко видоспецифичны. Именно они контролируют завершение акта спаривания и переноса сперматофора в женское половое отверстие (Allen, 1992).
У однохозяинного В. microplus самцы сразу после линьки отыскивают еще не перелинявших женских нимф и повторно присасываются рядом с ними. Спаривание происходит между питавшимися до этого несколько суток самцами и самками. Самки и нимфы этого вида выделяют 2,6-дихлорфенол, и вероятно, он совместно с другими компонентами выполняет функции полового аттрактанта (Bruyne, Guerin, 1994; Falk-Vairant et al., 1994).
У A. hebraeum и A. variegatum питающиеся самцы выделяют феромоны, привлекающие к ним голодных самцов, самок и нимф своего вида. Эти же феромоны стимулируют присасывание клещей в скоплениях на определенных частях тела хозяина. Описываемые феромоны скопления-прикрепления имеют в своем составе бензальдегид, 2,6-дихлорфенол, метилсалицилат, салицилальдегид и другие компоненты.
У I. ricinus и, видимо, у других видов этого рода фовеа с феромонными железами недоразвиты, но у них имеются аттрактантные феромоны. Последние выделяются голодными самцами и самками и привлекают клещей независимо от пола. Благодаря подобным феромонам возможно широкое распространение спаривания у голодных особей этого вида (Graf, 1978b).
Влияние спаривания на питание. Осеменение клещей является главным пусковым механизмом для нормального питания самок, переваривания ими пищи и оогенеза. Питание самок Amblyomminae без самцов вело к значительному удлинению сроков их нахождения на хозяине и невозможности достижения состояния нормального насыщения. В наших опытах (Балашов, 1967) средние сроки насыщения осемененных самок клещей Н. punctata, D. reticulatus, R. turanicus, H. asiaticum составляли в среднем 7—10 сут. Самки этих видов прикреплялись к хозяину и начинали питание и при отсутствии самцов. Однако их пребывание на теле хозяев затягивалось до 1—2 мес, и ни в одном случае они не достигали нормального насыщения. Даже наиболее упитанные особи весили по окончании питания значительно меньше, чем нормально осемененные. Обычно подобные неосемененные особи погибали недопитавшимися на теле хозяина или вскоре после его оставления.
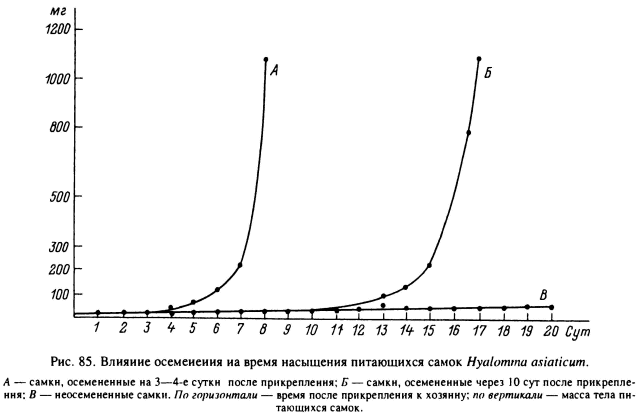
Первые 3—4 сут питание неосемененных самок протекало нормально и не отличалось от контроля (рис. 85). По достижении определенной степени насыщения дальнейшее увеличение массы, однако, прекращалось или шло в очень замедленном темпе. Этот переломный момент обычно наступал на 4—6-е сутки прикрепления по достижении массы 40—60 мг. Наоборот, у оплодотворенных особей в этот момент скорость питания заметно увеличилась, они вступали в период растяжения и отпадали через 1—3 сут. В случае продолжения питания неоплодотворенных особей у них отсутствовал четко выраженный период быстрого насыщения, а сроки отпадения с хозяина недопитавшихся клещей растягивались до нескольких недель. Если к подобным недопитавшимся самкам подсадить самцов того же вида, то через несколько дней происходит спаривание и затем быстрое насыщение..В опытах с подсадкой к подобным недопитавшимся самкам голодных самцов последним для приобретения способности к спариванию требовалось 3— 5 сут питания. После спаривания осемененные самки заканчивали питание за 3— 5 сут и отпадали. При этом масса их тела была близка к норме.
У девственных самок I. ricinus (рис. 84) первые 3—4 сут питание протекает как и у осемененных, и масса тела в обеих группах существенно не отличается. На 4—6-е сутки осемененные самки быстро набирают массу, заканчивают питание и отпадают. У девственных самок фаза быстрого насыщения отсутствовала. На протяжении последующих 10—15 сут масса их тела продолжала медленно увеличиваться, но даже на 15—20-е сутки питания она была в несколько раз меньше, чем у нормально напитавшихся, и не превышала 15—30 мг. В случае осеменения подобных самок они в течение следующих 3—5 сут завершали питание и, достигнув нормальной массы в 150—300 мг, отпадали с хозяина. Механизмом стимулирующего воздействия спаривания на нормальное завершение питания служит введение сперматофора во влагалище самки (Graf, 1978b).
Описанное явление торможения питания неоплодотворенных самок несомненно представляет приспособление для обеспечения встречи полов и оплодотворения. Шансы встречи малоподвижных и часто значительно удаленных друг от друга самцов и самок иксодид вне организма хозяина очень малы. Задержка неоплодотворенных недопитавшихся самок на прокормителях, как и длительное нахождение на их теле самцов — одна из своеобразных адаптации к пастбищному типу паразитизма.
Партеногенез. Откладка небольшого числа жизнеспособных партеногенетических яиц неоплодотворенными самками описана для клещей Н. anatolicum, H. marginatum, R. bursa, R. sanguineus (Первомайский, 1949), 2 видов рода Haemaphysalis (Жмаева, 1950; Лаптев, 1963) и В. microplus (Stone, 1963). В наших опытах неоплодотворенные недопитавшиеся самки Н. punctata, D. reticulatus и R. turanicus откладывали от 3 до 150 яиц. Лишь единичные из них развивались и давали личинок, тогда как большая часть вскоре высыхала. У популяции D. variabilis с одного из островов Атлантического побережья США обнаружена повышенная способность к партеногенезу и до стадии личинки развивалось от 5 до 50 % отложенных яиц (Nagar, 1967).
Партеногенез как нормальный способ размножения, по-видимому, свойствен немногим видам иксодид. Он обнаружен у обитающего в Юго-Восточной Азии A. cordiferum и A. rotundatum из Центральной и Южной Америки. Наиболее детально он изучен у Н. bispinosa, у которого обнаружены бисексуальная диплоидная и партеногенетическая триплоидная расы. Бисексуальная раса распространена в Приморье, Северо-Восточном Китае, Корее, островах Хонсю и Кюсю (Япония). Партеногенетическая раса встречается в Новой Зеландии, Австралии, на Хоккайдо и мелких островах вокруг Японии и Приморья. На одном из островов у побережья Кореи обнаружена также анеуплоидная раса, способная к половому размножению и партеногенезу (Oliver, 1989).
Подготовка к яйцекладке. Превращение кратковременного акта питания в продолжительную стадию развития на теле хозяина обеспечивает не только максимальное использование прокормителя как источника пищи, но и создает исключительно благоприятные условия для развития яиц. Рост яичников наблюдается с первых же часов питания. У напитавшихся и готовых к отпадению самок линейные размеры яичника увеличиваются в 10—12 раз и в них в небольшом числе содержатся готовые к оплодотворению ооциты (табл. XXXI, 1—3). После отпадения с хозяина продолжается развитие ооцитов и первая их порция готова к откладке спустя 1—2 нед. У неоплодотворенных самок рост яичников в период нахождения на хозяине происходит значительно медленнее, а у отпавших недопитавшихся особей они значительно меньше, чем после нормального насыщения. После отпадения с хозяина размеры яичников у подобных неоплодотворенных особей почти не увеличиваются и лишь немногие ооциты достигают нормальных размеров и выводятся в яйцеводы. Торможение яйцекладки у неоплодотворенных питающихся самок проявляется уже на стадии накопления желтка.
Сроки яйцекладки. Полностью напитавшаяся самка в зависимости от окружающей температуры приступает к яйцекладке через несколько дней или недель, а сама откладка занимает от 3 нед до 1—2 мес. Сроки наступления яйцекладки и ее продолжительность в первую очередь определяются окружающей температурой. В меньшей степени на эти показатели влияют относительная влажность и степень насыщения самок. Значительная задержка начала яйцекладки наблюдается при впадении напитавшихся самок в диапаузу.
У самок таежного клеща, по нашим наблюдениям, при температуре 20 °С и относительной влажности 95 % яйцекладка в среднем начиналась на 14-е сутки и продолжалась 36 сут, а при 26 °С — на 11-е сутки и занимала 28 сут. Более 3/4 яиц откладывалось за первую неделю яйцекладки. В природных местообитаниях этого вида в Карелии напитавшиеся в мае самки приступали к яйцекладке через 21—25 сут по отпадении, а напитавшиеся в конце июня—июле — через 10— 11 сут в зависимости от температурных условий (Хейсин и др., 1955). В Красноярском крае напитавшиеся в мае самки приступали к яйцекладке через 17— 25 сут, а в июле — через 8—15 сут (Таежный клещ..., 1985). Процесс яйцекладки в зависимости от температурного режима занимал от 1.5 до 2.5 мес.
У I. ricinus в зависимости от весенне-летнего или осеннего питания самок сроки подготовки к яйцекладке могут существенно различаться. В первом случае сроки начала откладки яиц в первую очередь зависят от окружающей температуры. В лабораторных условиях при 22 °С в мае—июне самки начинали откладывать яйца через 4—7 сут по отпадении, а при питании в сентябре—октябре — через 22 сут и более. Сроки яйцекладки варьировали от 1 до 2 мес (Honzakova, 1971). В природных местообитаниях в Ирландии самки весенних сроков питания приступали к яйцекладке через 14—21 сут и она продолжалась около 100 сут, а самки осеннего питания — через 21—28 сут и яйцекладка занимала более 200 сут (Gray, 1991). В условиях Карелии самки, отпавшие в начале мая, приступали к яйцекладке через 36—49 сут, напитавшиеся в июне—июле — через 19—24 сут, а часть самок августовского-сентябрьского питания не начиная яйцекладки уходила на зимовку (Хейсин и др., 1955).
У I. lividus в отличие от I. persulcatus и I. ricinus откладка яиц начинается через 3—8 сут после отпадения самок с птенцов и занимает 12—18 сут. Столь быстрые сроки развития становятся возможными благодаря обитанию клещей в гнездах ласточек-береговушек. Внутри гнезд в период яйцекладки клещей температура воздуха повышается до 30—40 °С за счет разогрева развивающимися птенцами (Глащинская-Бабенко, 1956).
У самок D. marginatus при бездиапаузном развитии при температуре 26—28 °С напитавшиеся особи приступали к яйцекладке через 5—7 сут и она занимала 15—20 сут. Наибольшее количество яиц было отложено на 2—3-й сутки с начала яйцекладки. В овражно-степных биотопах Воронежской области напитавшиеся в апреле самки начинали яйцекладку через 3—16 сут по отпадении с хозяина и она занимала 18—35 сут (Покровская, 1949). В Карагандинской области самки этого вида при весеннем насыщении приступали к яйцекладке на 7-е сутки после отпадения. При летнем и осеннем насыщении самки впадали в диапаузу и приступали к откладке яиц спустя 3—4 мес после отпадения. В природе подобные диапаузирующие особи уходили на зимовку и приступали к откладке яиц весной следующего года (Ламанова, 1962).
Прямая зависимость сроков яйцекладки от температуры отмечена для многих видов иксодид в пустынных районах Туркмении (Бердыев, 1980). Самки Н. asiaticum, отпавшие с хозяев в марте, приступали к яйцекладке через 41—59 сут, отпавшие в апреле — через 17—28 и в июне—июле — через 4—10 сут. Процесс яйцекладки за этот период сократился с 41 до 6 сут. Температура почвы в местообитаниях напитавшихся особей за период наблюдений повысилась с 15—16 до 26—34 °С. Сходное сокращение сроков подготовки к яйцекладке и откладки яиц отмечено в тех же условиях для самок более поздних сроков насыщения клещей Н. anatolicum, H. dromedarii, H. detritum и R. turanicus.
Яйцекладка. У напитавшихся и отпавших с хозяина самок во время подготовки к яйцекладке завершается развитие первой порции яиц и они поступают в полость яичника. Овуляция, по-видимому, обусловлена перистальтическими сокращениями стенок яичника и усилением внутреннего гидростатического давления при сокращении дорсовентральной мускулатуры идиосомы. В результате зрелые ооциты раздвигают клетки ножек и попадают в яйцеводы. Далее яйца передвигаются одно за другим по яйцеводам по направлению к влагалищу. Сами яйцеводы при этом приобретают характерную четкосидную форму вследствие растяжения их стенок яйцами. Оболочка яйца до его выхода из тела клеща остается мягкой и эластичной, что облегчает продвижение по сравнительно тонким яйцеводам. После откладки оболочка яйца сразу же затвердевает.
У яйцекладущих самок происходит характерное изменение формы тела. Задняя половина идиосомы сплющивается, а передняя образует горб. Гнатосома, располагающаяся у напитавшихся особей перпендикулярно главной оси тела, слегка подгибается вентрально. Навстречу ей в переднем направлении происходит смещение наружного полового отверстия.
Откладка яйца начинается с выворачивания вестибулярного отдела влагалища наружу с образованием короткого трубчатого яйцеклада. Одновременно хоботок подгибается вентрально и прижимается к идиосоме. Из дорсальной части камеростомальной складки выдвигаются рога органа Женэ. Они с боков прилегают к вывернутому наружу цервикальному отделу влагалища. Из последнего в этот момент выталкивается яйцо. Оно сразу же охватывается рогами органа Женэ и обволакивается его секретом. Затем гнатосома возвращается в свое первоначальное положение и яйцо переносится на переднедорсальную сторону идиосомы. В подобной же последовательности повторяется цикл откладки следующего яйца. Новые яйца склеиваются с уже отложенными, так что кладка одной самки представляет из себя единую компактную массу яиц.
У I. persulcatus процесс откладки одного яйца занимает при температуре 20— 22 °С 5—15 мин (Таежный клещ..., 1985), у I. ricinus — 3—12 (Lees, Beament, 1948), у D. marginatus при 28 °C — 1—5 (Hueli et al., 1988) и у Н. anatolicum — 2—8 мин (Snow, Arthur, 1966). Яйцекладка нормально напитавшихся самок в зависимости от их видовой принадлежности, окружающей температуры и в меньшей степени других факторов окружающей среды занимает от 2 нед до 1—2 мес. В наших лабораторных культурах при постоянной температуре 22—25 °С и оптимальной влажности у I. persulcatus яйцекладка продолжалась 16—34 сут, у I. ricinus — 19—31, у D. reticulatus — 11—24, у Н. concinna — 24—33, у R. turanicus — 15—33 и у Я. asiaticum — 22—39 сут. В указанные сроки наибольшая яйцевая продуктивность наблюдается в первые несколько дней яйцекладки, после чего ее темп существенно снижается.
В природных местообитаниях клещей значительные различия наблюдаются в продолжительности яйцекладки в разные сезоны года и в разных по теплообеспеченности биотопах. В условиях Карелии у I. persulcatus в июне—июле яйцекладка в более теплом лиственном лесу занимала 35—37 сут и в более прохладном смешанном лесу — 40—48 суг (Хейсин и др., 1955). В Туркмении в зависимости от яйцекладки весной или летом сроки ее варьировали у Н. asiaticum от 6 до 41 суг, у Н. dromedarii — от 8 до 60 и у R. turanicus — от 7 до 45 сут (Бердыев, 1980).
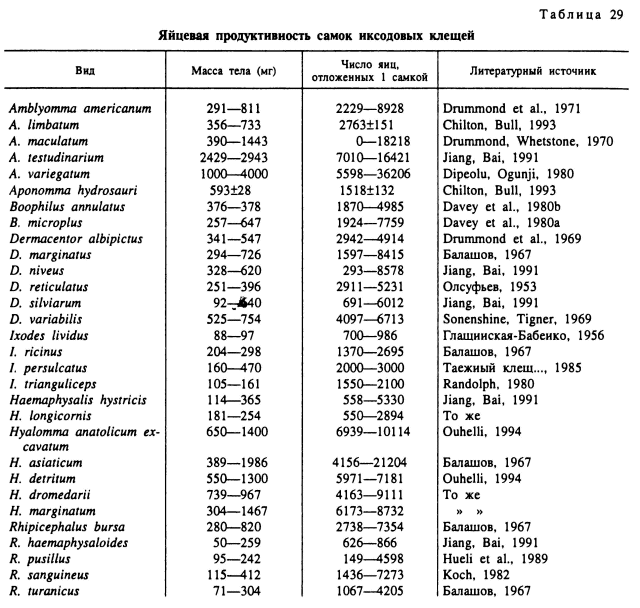
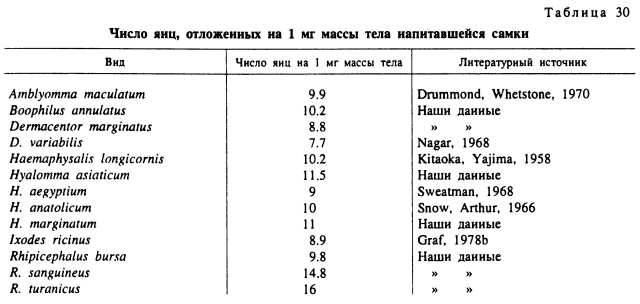
Яйцевая продуктивность. Полностью напитавшимся самкам иксодовых клещей принадлежат абсолютные рекорды плодовитости среди кровососущих членистоногих. Так, самки наиболее крупных видов (роды Hyalomma и Amblyomma) откладывают в среднем 15 000—20 000 яиц с максимумом в 21 204 для Н. asiaticum (Балашов, 1967) и 36 206 для A. variegatum (Dipeolu, Ogunji, 1980). Самки средних по размерам тела видов (Dermacentor, Rhipicephalus, Boophilus) откладывают 3000—6000 яиц и наиболее мелких норовых видов Ixodes и Haemaphysalis — около 1000 яиц (табл. 29).
У недопитавшихся оплодотворенных самок иксодид яйцекладка происходит только по достижении определенной минимальной массы тела, но количество оплодотворенных яиц бывает много меньше нормального. У В. microplus небольшое количество яиц откладывали самки, достигшие массы 17 мг и более (Bennett, 1974). Среди имеющих небольшие размеры тела I. ricinus и R. turanicus яйцекладка отмечена у самок массой не менее 20—30 мг, а у более крупных D. reticulatus, H. anatolicum, H. asiaticum — при массе не менее 45—73 мг (табл. 31). Из приведенных цифр видно, что яйцекладка возможна уже при достижении массы тела, близкой к 10 % от таковой нормально напитавшихся особей.
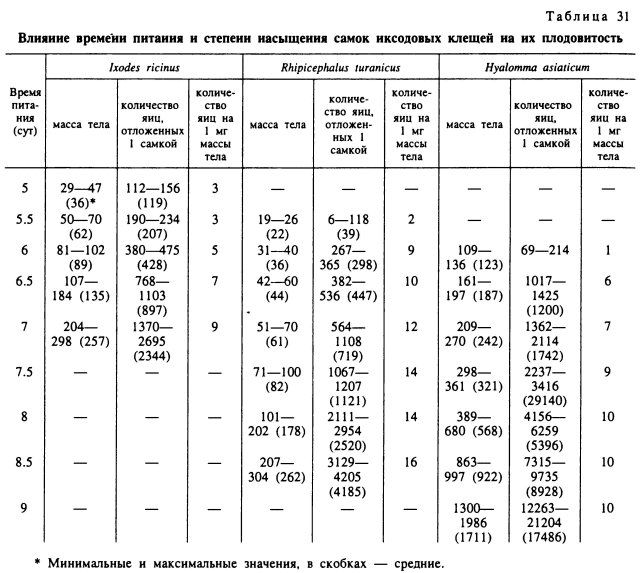
Количество откладываемых яиц с увеличением массы тела клещей обычно быстро возрастает как в абсолютных цифрах, так и в пересчете числа яиц на 1 мг массы тела самки. Последний показатель особенно важен, так как он отражает эффективность используемой при питании пищи. Максимальная яйцевая продуктивность на 1 мг массы достигается у полностью напитавшихся самок. Этот показатель в зависимости от видовых (Особенностей клещей и степени насыщения самок варьирует от 9 до 15 яиц на 1 мг массы тела (табл. 30). У недопитавшихся особей яйцевая продуктивность на единицу массы тела снижается в несколько раз (табл. 31). Например, у нормально напитавшихся самок I. ricinus, весящих 204— 298 мг, на 1 мг массы откладывалось 9 яиц, а у недопитавшихся особей массой 29—47 мг — всего 3 яйца. У недопитавшихся R. turanicus при массе 19—26 мг на 1 мг приходилось 2 яйца, а у нормально упитанных — 16. У напитавшихся Н. asiaticum яйцепродукция на 1 мг в 10 раз больше, чем у недопитавшихся.
Масса тела закончивших яйцекладку самок уменьшается в 3—4 раза по сравнению с таковой сразу по окончании питания. Например, у R. sanguineus она уменьшается на 75 % и у D. variabilis — на 74 % (Nagar, 1968). Большая часть этих потерь обусловлена переходом белков, липоидов и углеводов тела самки в откладываемые ею яйца, а меньшую часть составляют энергетические затраты. Так, у В. annulatus масса отложенных яиц составляет в среднем 61.6 % от массы тела свеженапитавшейся самки (Davey et al., 1980), у D. marginatus — 63.9 (Hueli et al., 1988), у A. americanum — 58.9 (Drummond et al., 1971) и у I. ricinus — 36.3—61.7 % (Graf, 1978b).
Наиболее полное использование поглощенной крови и тканей хозяина достигается у нормально напитавшихся особей. Например, у напитавшихся самок В. microplus и Н. bispinosa в отложенные яйца, главным образом в форме желтка, переходит 33.2—46.8 % белкового азота и 6.7—12.8 % железа (в виде гемопротеидов) от поглощенной крови хозяина (Kitaoka, 1961). При неблагоприятных внешних условиях эти показатели снижаются и большая доля питательных веществ идет на поддержание основного обмена веществ в организме. Так, при бездиапаузном развитии самок D. marginatus (яйцекладка через 7 сут после отпадения) на 1 клеща приходилось 6200 яиц, а при яйцекладке после 6-месячной диапаузы — только 2500 яиц (Ламанова, 1965). Особенно сильное снижение яйцевой продуктивности происходит вследствие питания клещей на резистентных к ним хозяевах, когда наблюдается общее уменьшение числа откладываемых яиц и их числа на 1 мг массы тела напитавшейся особи.
У голодных клещей яичники небольших размеров и независимо от возраста самок содержат только ооциты 1 и II стадий развития. После прикрепления к хозяину и начала питания процессы оогенеза возобновляются, но при отсутствии осеменения отложение желтка в ооцитах не наблюдается. Стимулом для вителлогенеза служит осеменение. Сразу же после него и параллельно со стадией быстрого насыщения клеща в кишечнике между обычными пищеварительными появляются желточные клетки. Размеры этих клеток быстро увеличиваются и достигают максимальных к началу яйцекладки. В процессе яйцекладки размеры и активность желточных клеток быстро убывают. Внутри желточных клеток и клеток жирового тела на последней стадии питания, при подготовке и во время яйцекладки выявляется в большом количестве белок — предшественник желтка, называемый вителлогенином. Из этих клеток вителлогенин поступает в гемолимфу, из нее путем эндоцитоза захватывается развивающимися ооцитами III и IV стадий развития.
Внутри ооцитов формируются многочисленные желточные гранулы, белок которых называют вителлином (Coons et al., 1989).
Процессы оогенеза и особенно отложения желтка в яйцах контролируются экдизоном. У голодных особей D. niveus и Н. longicornis содержание экдизона в организме было минимальным и существенно не менялось до начала питания. У питающихся особей первый небольшой пик экдизона приходился на период спаривания. После спаривания содержание экдизона в организме клещей продолжало расти и достигало максимума у напитавшихся самок перед отпадением. В гемолимфе и синганглии максимум концентрации экдизона отмечен перед началом яйцекладки. В яичнике содержание экдизона быстро увеличивалось через 3 сут после окончания питания и достигало максимума к началу яйцекладки. Часть экдизона переходила в отложенные яйца (Jiang Zaijie et al., 1994).
Значительные различия в яйцевой продуктивности на единицу массы в зависимости от степени упитанности объясняются особенностями развития ооцитов. У недопитавшихся особей начинает развиваться значительно больше яйцеклеток, чем могут закончить свой рост за счет поглощенных питательных веществ. Даже у самок, поглотивших пищи меньше, чем требуется для начала яйцекладки, наблюдается заметное увеличение размеров яичников, а многие ооциты находятся на разных стадиях заполнения желтком4. У недопитавшихся особей, способных к яйцекладке, размеры яичников и количество ооцитов, нормально закончивших развитие, значительно меньше, чем после нормального насыщения. При этом многие приступившие к отложению желтка ооциты так и не заканчивают роста из-за недостатка пищевых резервов. В то же время они поглощают много белков крови хозяина и вызывают общее снижение яйцевой продуктивности на 1 мг массы тела. Следует отметить, что чем выше степень насыщения клеща, тем в меньшем числе встречаются подобные (не закончившие развития) ооциты в яичниках самок после окончания яйцекладки.
В яичниках закончивших яйцекладку и погибающих самок, до этого нормально напитавшихся на хозяине, всегда сохраняется значительное количество не приступивших к росту ооцитов I—II стадий и небольшое число ооцитов III—IV стадий, не успевших закончить своего развития (табл. XXXI, 4). Последнее, вероятно, отражает утраченную иксодидами способность к прохождению нескольких гонотрофических циклов, которая была свойственна их примитивным предкам.
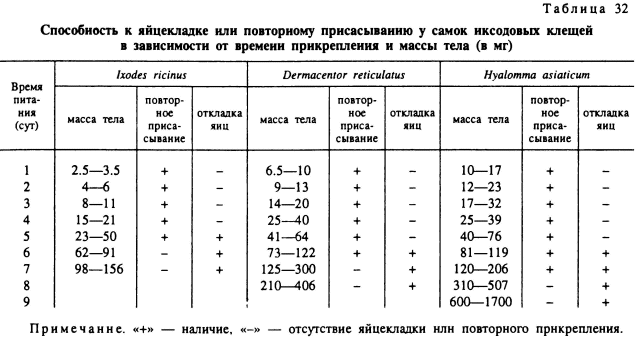
Повторное питание. Снижение числа отложенных яиц или отсутствие яйцекладки у недопитавшихся самок отчасти компенсируется их способностью к повторному прикреплению и окончанию питания на новом хозяине. В наших опытах (табл. 32) осторожно снятые с кролика недопитавшиеся самки 5 видов охотно прикреплялись к новому хозяину и нормально продолжали питание. Общая продолжительность питания, за вычетом времени перерыва, практически не отличалась от нормальной. Масса их тела после отпадения также заметно не отличалась от таковой нормально питавшихся клещей. С определенного момента недопитавшиеся самки способны как к повторному прикреплению к хозяину, так и к яйцекладке. Этот период обычно продолжается 1—2 сут. Затем способность к повторному прикреплению утрачивается, а число откладываемых яиц заметно увеличивается. Потеря способности к возобновлению питания после насильственного открепления клеща обычно совпадает с началом периода быстрого растяжения покровов тела. К этому времени самки I. ricinus и R. turanicus достигают массы не менее 60—75 мг, a D. reticulatus, H. anatolicum, H. asiaticum — 110—125 мг.
Способность к повторному прикреплению у насильственно удаленных с хозяина клещей сохраняется до их гибели. Однако их жизнеспособность значительно ниже, чем у не начинавших питаться голодных особей, и немногие из них выживают больше 1—2 мес. Резко различаются недопитавшиеся и нормально упитанные самки по своему поведению. У первых по удалении с хозяина сохраняется значительная подвижность и комплекс реакций, связанных с подстереганием хозяина (подъем на растительность, поза ожидания и др.). Напротив, у нормально напитавшихся и недопитавшихся самок, способных к откладке и утративших способность к повторному питанию, хорошо выражены реакции положительного геотропизма и отрицательного фототаксиса, и они стремятся укрыться в щелях, трещинах и других убежищах. Инстинкт нападения на хозяев у них полностью утрачен.
РАЗМНОЖЕНИЕ
Самкам иксодовых клещей свойственна жесткая гонотрофическая гармония, так как необратимые морфофизиологические изменения в их организме во время питания и подготовки к яйцекладке делают невозможным прохождение повторных гонотрофических циклов. Нормальное завершение гонотрофического цикла связано с успешным насыщением, которое возможно только у осемененных самок.
Встреча полов. Самки I. ricinus, I. persulcatus (Балашов, 1967; Graf, 1978b), I. scapularis (Yuval, Spielman, 1990), I. rubicundus (Fourie, Kok, 1995), I. trianguliceps (Randolph, 1980) и, вероятно, других видов рода Ixodes способны к копуляции в голодном состоянии. Это неоднократно наблюдали как в природе, так и в лабораторных культурах клещей. В природных популяциях I. persulcatus среди самок, собранных с растительности, по данным вскрытий, осемененные особи составляли от 0 до 50—65 %. При этом за сезон активности клещей доля осемененных самок постепенно увеличивается. Этот же показатель возрастал при увеличении плотности клещей и особенно обилия активных особей (Успенский, Репкина, 1978; Бабенко и др., 1979). У I. ricinus доля осемененных самок, собранных с растительности в Швейцарии, варьировала от 27 до 100 % (Graf, 1978b) и в Латвии — 22—30 % (Бабенко и др., 1979). Высокий процент осемененных самок выявлен также среди активных голодных особей I. rubicundus и I. scapularis. На хозяев нападают осемененные и девственные самки видов этого рода, а также самцы. Поэтому осеменение девственных самок возможно и на теле хозяина. В местах присасывания клещей в большом количестве встречаются самцы, копулирующие с питающимися самками.
У видов подсем. Amblyomminae голодные самцы и самки не способны к копуляции. Исключение, возможно, составляют немногие виды Аропотта и Amblyomma, у которых зрелые сперматиды обнаружены в половых путях не питавшихся самцов (Oliver, 1989). У остальных видов голодные самцы не способны к спариванию. Зрелые сперматиды появляются у них на 3—5-е сутки с начала питания. После этого они приступают к копуляции и осеменяют питающихся самок. Голодные самки Amblyomminae также лишены способности к спариванию и осеменению.
У I. ricinus максимальное число копуляций у одного самца было 4, но чаще их бывает только 2. Затем подобные самцы быстро погибают. В то же время девственные самцы живут в благоприятных условиях до года и более (Graf, 1978b). Самцы Amblyomminae способны к значительно большему числу повторных спариваний. В промежутках между спариваниями они повторно прикрепляются к хозяину, питаются и у них возобновляются процессы сперматогенеза. Благодаря подобному восполнению половых продуктов один самец может последовательно осеменить несколько десятков самок. В литературе описано осеменение самцом Н. marginatum 25 самок (Первомайский, 1954), самцами A. americanum — до 27— 37 самок за 68—127 сут (Gladney, Drummond, 1970) и до 22 самок D. variabilis (Oliver, 1982).
Спаривание. Наиболее полно процесс спаривания описан у I. ricinus (Graf, 1978b). После установления самцом контакта с самкой он закрепляется на ее вентральной стороне при помощи II и III пар ног, обеспечивая тесный контакт между вентральными поверхностями партнеров. Ноги самки не участвуют в копуляции, и спаривающиеся самки свободно перемещаются. После закрепления на теле самки самец подводит свои ротовые органы к генитальному отверстию и погружает в него гипостом и хелицеры. Хоботок клеща при этом располагается перпендикулярно к спинному щитку, а пальпы остаются снаружи и лишь раздвигаются плоско в стороны от полового отверстия. В этом положении самец может оставаться несколько часов или даже суток (рис. 100, А).
Второй этап копуляции начинается с повторяющихся расширяющих движений хоботка в половом отверстии самки, сопровождающихся выделением капелек слюны. Затем из мужского полового отверстия выходит наружу эктосперматофор, размеры которого быстро увеличиваются за счет заполнения половыми продуктами (рис. 100, Б, В). После этого хоботок выдвигается из полового отверстия самки и подгибается на вентральную сторону, так что хелицеры захватывают шейку сперматофора и переносят его снаружи к женскому половому отверстию (рис. 100, Г, Д). При переносе сперматофора на дистальных концах хелицер видны капельки жидкости, вероятно являющейся секретом слюнных желез. С помощью пальцев хелицер шейка сперматофора вводится внутрь полового отверстия (рис. 100, Е). По завершении копуляции самец оставляет самку.
Содержимое эктосперматофора переливается в эндосперматофор, находящийся уже внутри центральной части влагалища. Спустя 48—72 ч после конца копуляции капсула эндосперматофора открывается в своей апикальной области и спермиофоры заполняют матку, размеры которой при этом сильно увеличиваются (рис. 101). Пустой эктосперматофор может оставаться прикрепленным к женскому половому отверстию в течение нескольких часов или дней по окончании спаривания.
У Amblyomminae процесс спаривания в основных чертах сходен с описанным выше для Ixodes, но имеются и определенные различия. Перед спариванием питающийся самец временно открепляется от хозяина в ответ на половые аттрактанты находящейся вблизи самки. В процессе спаривания удержание самцов обеспечивается взаимным зацеплением ног обоих партнеров. В половое отверстие вводятся только пальцы хелицер, а гипостом остается снаружи вместе с пальпами. У некоторых видов (Н. excavatum, R. sanguineus) перенос сперматофора осуществляется скользящими движениями всей идиосомы самца и хелицеры на это время не выводятся из полового отверстия самки. У других видов (A. variegatum, H. marginatum) перенос сперматофора, как и у Ixodes, осуществляется только с помощью ротовых частей (Feldman-Muhsam, Borut, 1983a).
Феромоны. Встреча полов и спаривание у иксодовых клещей регулируются особыми химическими веществами — феромонами. Они выделяются во время питания особями одного или обоих полов, стимулируют соответствующие рецепторы и вызывают групповое прикрепление самцов и самок одного вида на определенных участках тела хозяина.
У видов родов Dermacentor, Hyalomma и Rhipicephalus питающиеся самки выцеляют летучие половые феромоны, главным компонентом которых у всех видов является 2,6-дихлорфенол (Sonenshine, 1985). Местом выделения этого типа феромонов служат особые железы, располагающиеся на аллоскутуме под фовеа. Они начинают функционировать только через несколько суток питания, и их активность совпадает с повышением содержания экдизона в организме. У голодных самок выделение полового феромона отсутствует. Нет реакции на них и у голодных самцов.
Питающиеся самцы воспринимают 2,6-дихлорфенол хеморецепторными сенсиллами передней ямки органа Галлера (табл. XXVI, 3). В ответ на их раздражение питающиеся самцы открепляются и двигаются в направлении питающихся самок. После установления тактильного контакта с последними самцы с готовыми половыми продуктами приступают к спариванию и осеменению самок. Самцы, не достигшие способности к осеменению, повторно прикрепляются к хозяину рядом с питающейся самкой и приступают к копуляции спустя несколько часов или суток по завершении созревания половых продуктов.
2,6-дихлорфенол не обладает видовой специфичностью, и собственно копуляция стимулируется другой группой уже видоспецифичных феромонов. Важная роль в их восприятии принадлежит группе хеморецепторных сенсилл у основания коготка I пары ног (Sonenshine, 1991). В процессе спаривания самцы контактируют пальцами хелицер с половым отверстием самки и окружающей его кутикулой. На этой стадии хеморецепторы пальцев хелицер воспринимают еще одну группу половых феромонов. Последние вырабатываются передними отделами женской половой системы, мало летучи и находятся на поверхности кутикулы. Одним из их главных компонентов являются насыщенные жирные кислоты с цепью из 14—22 атомов углерода. Феромоны этой группы высоко видоспецифичны. Именно они контролируют завершение акта спаривания и переноса сперматофора в женское половое отверстие (Allen, 1992).
У однохозяинного В. microplus самцы сразу после линьки отыскивают еще не перелинявших женских нимф и повторно присасываются рядом с ними. Спаривание происходит между питавшимися до этого несколько суток самцами и самками. Самки и нимфы этого вида выделяют 2,6-дихлорфенол, и вероятно, он совместно с другими компонентами выполняет функции полового аттрактанта (Bruyne, Guerin, 1994; Falk-Vairant et al., 1994).
У A. hebraeum и A. variegatum питающиеся самцы выделяют феромоны, привлекающие к ним голодных самцов, самок и нимф своего вида. Эти же феромоны стимулируют присасывание клещей в скоплениях на определенных частях тела хозяина. Описываемые феромоны скопления-прикрепления имеют в своем составе бензальдегид, 2,6-дихлорфенол, метилсалицилат, салицилальдегид и другие компоненты.
У I. ricinus и, видимо, у других видов этого рода фовеа с феромонными железами недоразвиты, но у них имеются аттрактантные феромоны. Последние выделяются голодными самцами и самками и привлекают клещей независимо от пола. Благодаря подобным феромонам возможно широкое распространение спаривания у голодных особей этого вида (Graf, 1978b).
Влияние спаривания на питание. Осеменение клещей является главным пусковым механизмом для нормального питания самок, переваривания ими пищи и оогенеза. Питание самок Amblyomminae без самцов вело к значительному удлинению сроков их нахождения на хозяине и невозможности достижения состояния нормального насыщения. В наших опытах (Балашов, 1967) средние сроки насыщения осемененных самок клещей Н. punctata, D. reticulatus, R. turanicus, H. asiaticum составляли в среднем 7—10 сут. Самки этих видов прикреплялись к хозяину и начинали питание и при отсутствии самцов. Однако их пребывание на теле хозяев затягивалось до 1—2 мес, и ни в одном случае они не достигали нормального насыщения. Даже наиболее упитанные особи весили по окончании питания значительно меньше, чем нормально осемененные. Обычно подобные неосемененные особи погибали недопитавшимися на теле хозяина или вскоре после его оставления.
Первые 3—4 сут питание неосемененных самок протекало нормально и не отличалось от контроля (рис. 85). По достижении определенной степени насыщения дальнейшее увеличение массы, однако, прекращалось или шло в очень замедленном темпе. Этот переломный момент обычно наступал на 4—6-е сутки прикрепления по достижении массы 40—60 мг. Наоборот, у оплодотворенных особей в этот момент скорость питания заметно увеличилась, они вступали в период растяжения и отпадали через 1—3 сут. В случае продолжения питания неоплодотворенных особей у них отсутствовал четко выраженный период быстрого насыщения, а сроки отпадения с хозяина недопитавшихся клещей растягивались до нескольких недель. Если к подобным недопитавшимся самкам подсадить самцов того же вида, то через несколько дней происходит спаривание и затем быстрое насыщение..В опытах с подсадкой к подобным недопитавшимся самкам голодных самцов последним для приобретения способности к спариванию требовалось 3— 5 сут питания. После спаривания осемененные самки заканчивали питание за 3— 5 сут и отпадали. При этом масса их тела была близка к норме.
У девственных самок I. ricinus (рис. 84) первые 3—4 сут питание протекает как и у осемененных, и масса тела в обеих группах существенно не отличается. На 4—6-е сутки осемененные самки быстро набирают массу, заканчивают питание и отпадают. У девственных самок фаза быстрого насыщения отсутствовала. На протяжении последующих 10—15 сут масса их тела продолжала медленно увеличиваться, но даже на 15—20-е сутки питания она была в несколько раз меньше, чем у нормально напитавшихся, и не превышала 15—30 мг. В случае осеменения подобных самок они в течение следующих 3—5 сут завершали питание и, достигнув нормальной массы в 150—300 мг, отпадали с хозяина. Механизмом стимулирующего воздействия спаривания на нормальное завершение питания служит введение сперматофора во влагалище самки (Graf, 1978b).
Описанное явление торможения питания неоплодотворенных самок несомненно представляет приспособление для обеспечения встречи полов и оплодотворения. Шансы встречи малоподвижных и часто значительно удаленных друг от друга самцов и самок иксодид вне организма хозяина очень малы. Задержка неоплодотворенных недопитавшихся самок на прокормителях, как и длительное нахождение на их теле самцов — одна из своеобразных адаптации к пастбищному типу паразитизма.
Партеногенез. Откладка небольшого числа жизнеспособных партеногенетических яиц неоплодотворенными самками описана для клещей Н. anatolicum, H. marginatum, R. bursa, R. sanguineus (Первомайский, 1949), 2 видов рода Haemaphysalis (Жмаева, 1950; Лаптев, 1963) и В. microplus (Stone, 1963). В наших опытах неоплодотворенные недопитавшиеся самки Н. punctata, D. reticulatus и R. turanicus откладывали от 3 до 150 яиц. Лишь единичные из них развивались и давали личинок, тогда как большая часть вскоре высыхала. У популяции D. variabilis с одного из островов Атлантического побережья США обнаружена повышенная способность к партеногенезу и до стадии личинки развивалось от 5 до 50 % отложенных яиц (Nagar, 1967).
Партеногенез как нормальный способ размножения, по-видимому, свойствен немногим видам иксодид. Он обнаружен у обитающего в Юго-Восточной Азии A. cordiferum и A. rotundatum из Центральной и Южной Америки. Наиболее детально он изучен у Н. bispinosa, у которого обнаружены бисексуальная диплоидная и партеногенетическая триплоидная расы. Бисексуальная раса распространена в Приморье, Северо-Восточном Китае, Корее, островах Хонсю и Кюсю (Япония). Партеногенетическая раса встречается в Новой Зеландии, Австралии, на Хоккайдо и мелких островах вокруг Японии и Приморья. На одном из островов у побережья Кореи обнаружена также анеуплоидная раса, способная к половому размножению и партеногенезу (Oliver, 1989).
Подготовка к яйцекладке. Превращение кратковременного акта питания в продолжительную стадию развития на теле хозяина обеспечивает не только максимальное использование прокормителя как источника пищи, но и создает исключительно благоприятные условия для развития яиц. Рост яичников наблюдается с первых же часов питания. У напитавшихся и готовых к отпадению самок линейные размеры яичника увеличиваются в 10—12 раз и в них в небольшом числе содержатся готовые к оплодотворению ооциты (табл. XXXI, 1—3). После отпадения с хозяина продолжается развитие ооцитов и первая их порция готова к откладке спустя 1—2 нед. У неоплодотворенных самок рост яичников в период нахождения на хозяине происходит значительно медленнее, а у отпавших недопитавшихся особей они значительно меньше, чем после нормального насыщения. После отпадения с хозяина размеры яичников у подобных неоплодотворенных особей почти не увеличиваются и лишь немногие ооциты достигают нормальных размеров и выводятся в яйцеводы. Торможение яйцекладки у неоплодотворенных питающихся самок проявляется уже на стадии накопления желтка.
Сроки яйцекладки. Полностью напитавшаяся самка в зависимости от окружающей температуры приступает к яйцекладке через несколько дней или недель, а сама откладка занимает от 3 нед до 1—2 мес. Сроки наступления яйцекладки и ее продолжительность в первую очередь определяются окружающей температурой. В меньшей степени на эти показатели влияют относительная влажность и степень насыщения самок. Значительная задержка начала яйцекладки наблюдается при впадении напитавшихся самок в диапаузу.
У самок таежного клеща, по нашим наблюдениям, при температуре 20 °С и относительной влажности 95 % яйцекладка в среднем начиналась на 14-е сутки и продолжалась 36 сут, а при 26 °С — на 11-е сутки и занимала 28 сут. Более 3/4 яиц откладывалось за первую неделю яйцекладки. В природных местообитаниях этого вида в Карелии напитавшиеся в мае самки приступали к яйцекладке через 21—25 сут по отпадении, а напитавшиеся в конце июня—июле — через 10— 11 сут в зависимости от температурных условий (Хейсин и др., 1955). В Красноярском крае напитавшиеся в мае самки приступали к яйцекладке через 17— 25 сут, а в июле — через 8—15 сут (Таежный клещ..., 1985). Процесс яйцекладки в зависимости от температурного режима занимал от 1.5 до 2.5 мес.
У I. ricinus в зависимости от весенне-летнего или осеннего питания самок сроки подготовки к яйцекладке могут существенно различаться. В первом случае сроки начала откладки яиц в первую очередь зависят от окружающей температуры. В лабораторных условиях при 22 °С в мае—июне самки начинали откладывать яйца через 4—7 сут по отпадении, а при питании в сентябре—октябре — через 22 сут и более. Сроки яйцекладки варьировали от 1 до 2 мес (Honzakova, 1971). В природных местообитаниях в Ирландии самки весенних сроков питания приступали к яйцекладке через 14—21 сут и она продолжалась около 100 сут, а самки осеннего питания — через 21—28 сут и яйцекладка занимала более 200 сут (Gray, 1991). В условиях Карелии самки, отпавшие в начале мая, приступали к яйцекладке через 36—49 сут, напитавшиеся в июне—июле — через 19—24 сут, а часть самок августовского-сентябрьского питания не начиная яйцекладки уходила на зимовку (Хейсин и др., 1955).
У I. lividus в отличие от I. persulcatus и I. ricinus откладка яиц начинается через 3—8 сут после отпадения самок с птенцов и занимает 12—18 сут. Столь быстрые сроки развития становятся возможными благодаря обитанию клещей в гнездах ласточек-береговушек. Внутри гнезд в период яйцекладки клещей температура воздуха повышается до 30—40 °С за счет разогрева развивающимися птенцами (Глащинская-Бабенко, 1956).
У самок D. marginatus при бездиапаузном развитии при температуре 26—28 °С напитавшиеся особи приступали к яйцекладке через 5—7 сут и она занимала 15—20 сут. Наибольшее количество яиц было отложено на 2—3-й сутки с начала яйцекладки. В овражно-степных биотопах Воронежской области напитавшиеся в апреле самки начинали яйцекладку через 3—16 сут по отпадении с хозяина и она занимала 18—35 сут (Покровская, 1949). В Карагандинской области самки этого вида при весеннем насыщении приступали к яйцекладке на 7-е сутки после отпадения. При летнем и осеннем насыщении самки впадали в диапаузу и приступали к откладке яиц спустя 3—4 мес после отпадения. В природе подобные диапаузирующие особи уходили на зимовку и приступали к откладке яиц весной следующего года (Ламанова, 1962).
Прямая зависимость сроков яйцекладки от температуры отмечена для многих видов иксодид в пустынных районах Туркмении (Бердыев, 1980). Самки Н. asiaticum, отпавшие с хозяев в марте, приступали к яйцекладке через 41—59 сут, отпавшие в апреле — через 17—28 и в июне—июле — через 4—10 сут. Процесс яйцекладки за этот период сократился с 41 до 6 сут. Температура почвы в местообитаниях напитавшихся особей за период наблюдений повысилась с 15—16 до 26—34 °С. Сходное сокращение сроков подготовки к яйцекладке и откладки яиц отмечено в тех же условиях для самок более поздних сроков насыщения клещей Н. anatolicum, H. dromedarii, H. detritum и R. turanicus.
Яйцекладка. У напитавшихся и отпавших с хозяина самок во время подготовки к яйцекладке завершается развитие первой порции яиц и они поступают в полость яичника. Овуляция, по-видимому, обусловлена перистальтическими сокращениями стенок яичника и усилением внутреннего гидростатического давления при сокращении дорсовентральной мускулатуры идиосомы. В результате зрелые ооциты раздвигают клетки ножек и попадают в яйцеводы. Далее яйца передвигаются одно за другим по яйцеводам по направлению к влагалищу. Сами яйцеводы при этом приобретают характерную четкосидную форму вследствие растяжения их стенок яйцами. Оболочка яйца до его выхода из тела клеща остается мягкой и эластичной, что облегчает продвижение по сравнительно тонким яйцеводам. После откладки оболочка яйца сразу же затвердевает.
У яйцекладущих самок происходит характерное изменение формы тела. Задняя половина идиосомы сплющивается, а передняя образует горб. Гнатосома, располагающаяся у напитавшихся особей перпендикулярно главной оси тела, слегка подгибается вентрально. Навстречу ей в переднем направлении происходит смещение наружного полового отверстия.
Откладка яйца начинается с выворачивания вестибулярного отдела влагалища наружу с образованием короткого трубчатого яйцеклада. Одновременно хоботок подгибается вентрально и прижимается к идиосоме. Из дорсальной части камеростомальной складки выдвигаются рога органа Женэ. Они с боков прилегают к вывернутому наружу цервикальному отделу влагалища. Из последнего в этот момент выталкивается яйцо. Оно сразу же охватывается рогами органа Женэ и обволакивается его секретом. Затем гнатосома возвращается в свое первоначальное положение и яйцо переносится на переднедорсальную сторону идиосомы. В подобной же последовательности повторяется цикл откладки следующего яйца. Новые яйца склеиваются с уже отложенными, так что кладка одной самки представляет из себя единую компактную массу яиц.
У I. persulcatus процесс откладки одного яйца занимает при температуре 20— 22 °С 5—15 мин (Таежный клещ..., 1985), у I. ricinus — 3—12 (Lees, Beament, 1948), у D. marginatus при 28 °C — 1—5 (Hueli et al., 1988) и у Н. anatolicum — 2—8 мин (Snow, Arthur, 1966). Яйцекладка нормально напитавшихся самок в зависимости от их видовой принадлежности, окружающей температуры и в меньшей степени других факторов окружающей среды занимает от 2 нед до 1—2 мес. В наших лабораторных культурах при постоянной температуре 22—25 °С и оптимальной влажности у I. persulcatus яйцекладка продолжалась 16—34 сут, у I. ricinus — 19—31, у D. reticulatus — 11—24, у Н. concinna — 24—33, у R. turanicus — 15—33 и у Я. asiaticum — 22—39 сут. В указанные сроки наибольшая яйцевая продуктивность наблюдается в первые несколько дней яйцекладки, после чего ее темп существенно снижается.
В природных местообитаниях клещей значительные различия наблюдаются в продолжительности яйцекладки в разные сезоны года и в разных по теплообеспеченности биотопах. В условиях Карелии у I. persulcatus в июне—июле яйцекладка в более теплом лиственном лесу занимала 35—37 сут и в более прохладном смешанном лесу — 40—48 суг (Хейсин и др., 1955). В Туркмении в зависимости от яйцекладки весной или летом сроки ее варьировали у Н. asiaticum от 6 до 41 суг, у Н. dromedarii — от 8 до 60 и у R. turanicus — от 7 до 45 сут (Бердыев, 1980).
Яйцевая продуктивность. Полностью напитавшимся самкам иксодовых клещей принадлежат абсолютные рекорды плодовитости среди кровососущих членистоногих. Так, самки наиболее крупных видов (роды Hyalomma и Amblyomma) откладывают в среднем 15 000—20 000 яиц с максимумом в 21 204 для Н. asiaticum (Балашов, 1967) и 36 206 для A. variegatum (Dipeolu, Ogunji, 1980). Самки средних по размерам тела видов (Dermacentor, Rhipicephalus, Boophilus) откладывают 3000—6000 яиц и наиболее мелких норовых видов Ixodes и Haemaphysalis — около 1000 яиц (табл. 29).
У недопитавшихся оплодотворенных самок иксодид яйцекладка происходит только по достижении определенной минимальной массы тела, но количество оплодотворенных яиц бывает много меньше нормального. У В. microplus небольшое количество яиц откладывали самки, достигшие массы 17 мг и более (Bennett, 1974). Среди имеющих небольшие размеры тела I. ricinus и R. turanicus яйцекладка отмечена у самок массой не менее 20—30 мг, а у более крупных D. reticulatus, H. anatolicum, H. asiaticum — при массе не менее 45—73 мг (табл. 31). Из приведенных цифр видно, что яйцекладка возможна уже при достижении массы тела, близкой к 10 % от таковой нормально напитавшихся особей.
Количество откладываемых яиц с увеличением массы тела клещей обычно быстро возрастает как в абсолютных цифрах, так и в пересчете числа яиц на 1 мг массы тела самки. Последний показатель особенно важен, так как он отражает эффективность используемой при питании пищи. Максимальная яйцевая продуктивность на 1 мг массы достигается у полностью напитавшихся самок. Этот показатель в зависимости от видовых (Особенностей клещей и степени насыщения самок варьирует от 9 до 15 яиц на 1 мг массы тела (табл. 30). У недопитавшихся особей яйцевая продуктивность на единицу массы тела снижается в несколько раз (табл. 31). Например, у нормально напитавшихся самок I. ricinus, весящих 204— 298 мг, на 1 мг массы откладывалось 9 яиц, а у недопитавшихся особей массой 29—47 мг — всего 3 яйца. У недопитавшихся R. turanicus при массе 19—26 мг на 1 мг приходилось 2 яйца, а у нормально упитанных — 16. У напитавшихся Н. asiaticum яйцепродукция на 1 мг в 10 раз больше, чем у недопитавшихся.
Масса тела закончивших яйцекладку самок уменьшается в 3—4 раза по сравнению с таковой сразу по окончании питания. Например, у R. sanguineus она уменьшается на 75 % и у D. variabilis — на 74 % (Nagar, 1968). Большая часть этих потерь обусловлена переходом белков, липоидов и углеводов тела самки в откладываемые ею яйца, а меньшую часть составляют энергетические затраты. Так, у В. annulatus масса отложенных яиц составляет в среднем 61.6 % от массы тела свеженапитавшейся самки (Davey et al., 1980), у D. marginatus — 63.9 (Hueli et al., 1988), у A. americanum — 58.9 (Drummond et al., 1971) и у I. ricinus — 36.3—61.7 % (Graf, 1978b).
Наиболее полное использование поглощенной крови и тканей хозяина достигается у нормально напитавшихся особей. Например, у напитавшихся самок В. microplus и Н. bispinosa в отложенные яйца, главным образом в форме желтка, переходит 33.2—46.8 % белкового азота и 6.7—12.8 % железа (в виде гемопротеидов) от поглощенной крови хозяина (Kitaoka, 1961). При неблагоприятных внешних условиях эти показатели снижаются и большая доля питательных веществ идет на поддержание основного обмена веществ в организме. Так, при бездиапаузном развитии самок D. marginatus (яйцекладка через 7 сут после отпадения) на 1 клеща приходилось 6200 яиц, а при яйцекладке после 6-месячной диапаузы — только 2500 яиц (Ламанова, 1965). Особенно сильное снижение яйцевой продуктивности происходит вследствие питания клещей на резистентных к ним хозяевах, когда наблюдается общее уменьшение числа откладываемых яиц и их числа на 1 мг массы тела напитавшейся особи.
У голодных клещей яичники небольших размеров и независимо от возраста самок содержат только ооциты 1 и II стадий развития. После прикрепления к хозяину и начала питания процессы оогенеза возобновляются, но при отсутствии осеменения отложение желтка в ооцитах не наблюдается. Стимулом для вителлогенеза служит осеменение. Сразу же после него и параллельно со стадией быстрого насыщения клеща в кишечнике между обычными пищеварительными появляются желточные клетки. Размеры этих клеток быстро увеличиваются и достигают максимальных к началу яйцекладки. В процессе яйцекладки размеры и активность желточных клеток быстро убывают. Внутри желточных клеток и клеток жирового тела на последней стадии питания, при подготовке и во время яйцекладки выявляется в большом количестве белок — предшественник желтка, называемый вителлогенином. Из этих клеток вителлогенин поступает в гемолимфу, из нее путем эндоцитоза захватывается развивающимися ооцитами III и IV стадий развития.
Внутри ооцитов формируются многочисленные желточные гранулы, белок которых называют вителлином (Coons et al., 1989).
Процессы оогенеза и особенно отложения желтка в яйцах контролируются экдизоном. У голодных особей D. niveus и Н. longicornis содержание экдизона в организме было минимальным и существенно не менялось до начала питания. У питающихся особей первый небольшой пик экдизона приходился на период спаривания. После спаривания содержание экдизона в организме клещей продолжало расти и достигало максимума у напитавшихся самок перед отпадением. В гемолимфе и синганглии максимум концентрации экдизона отмечен перед началом яйцекладки. В яичнике содержание экдизона быстро увеличивалось через 3 сут после окончания питания и достигало максимума к началу яйцекладки. Часть экдизона переходила в отложенные яйца (Jiang Zaijie et al., 1994).
Значительные различия в яйцевой продуктивности на единицу массы в зависимости от степени упитанности объясняются особенностями развития ооцитов. У недопитавшихся особей начинает развиваться значительно больше яйцеклеток, чем могут закончить свой рост за счет поглощенных питательных веществ. Даже у самок, поглотивших пищи меньше, чем требуется для начала яйцекладки, наблюдается заметное увеличение размеров яичников, а многие ооциты находятся на разных стадиях заполнения желтком4. У недопитавшихся особей, способных к яйцекладке, размеры яичников и количество ооцитов, нормально закончивших развитие, значительно меньше, чем после нормального насыщения. При этом многие приступившие к отложению желтка ооциты так и не заканчивают роста из-за недостатка пищевых резервов. В то же время они поглощают много белков крови хозяина и вызывают общее снижение яйцевой продуктивности на 1 мг массы тела. Следует отметить, что чем выше степень насыщения клеща, тем в меньшем числе встречаются подобные (не закончившие развития) ооциты в яичниках самок после окончания яйцекладки.
В яичниках закончивших яйцекладку и погибающих самок, до этого нормально напитавшихся на хозяине, всегда сохраняется значительное количество не приступивших к росту ооцитов I—II стадий и небольшое число ооцитов III—IV стадий, не успевших закончить своего развития (табл. XXXI, 4). Последнее, вероятно, отражает утраченную иксодидами способность к прохождению нескольких гонотрофических циклов, которая была свойственна их примитивным предкам.
Повторное питание. Снижение числа отложенных яиц или отсутствие яйцекладки у недопитавшихся самок отчасти компенсируется их способностью к повторному прикреплению и окончанию питания на новом хозяине. В наших опытах (табл. 32) осторожно снятые с кролика недопитавшиеся самки 5 видов охотно прикреплялись к новому хозяину и нормально продолжали питание. Общая продолжительность питания, за вычетом времени перерыва, практически не отличалась от нормальной. Масса их тела после отпадения также заметно не отличалась от таковой нормально питавшихся клещей. С определенного момента недопитавшиеся самки способны как к повторному прикреплению к хозяину, так и к яйцекладке. Этот период обычно продолжается 1—2 сут. Затем способность к повторному прикреплению утрачивается, а число откладываемых яиц заметно увеличивается. Потеря способности к возобновлению питания после насильственного открепления клеща обычно совпадает с началом периода быстрого растяжения покровов тела. К этому времени самки I. ricinus и R. turanicus достигают массы не менее 60—75 мг, a D. reticulatus, H. anatolicum, H. asiaticum — 110—125 мг.
Способность к повторному прикреплению у насильственно удаленных с хозяина клещей сохраняется до их гибели. Однако их жизнеспособность значительно ниже, чем у не начинавших питаться голодных особей, и немногие из них выживают больше 1—2 мес. Резко различаются недопитавшиеся и нормально упитанные самки по своему поведению. У первых по удалении с хозяина сохраняется значительная подвижность и комплекс реакций, связанных с подстереганием хозяина (подъем на растительность, поза ожидания и др.). Напротив, у нормально напитавшихся и недопитавшихся самок, способных к откладке и утративших способность к повторному питанию, хорошо выражены реакции положительного геотропизма и отрицательного фототаксиса, и они стремятся укрыться в щелях, трещинах и других убежищах. Инстинкт нападения на хозяев у них полностью утрачен.