Иксодовые клещи — паразиты и переносчики инфекций
Органы чувств
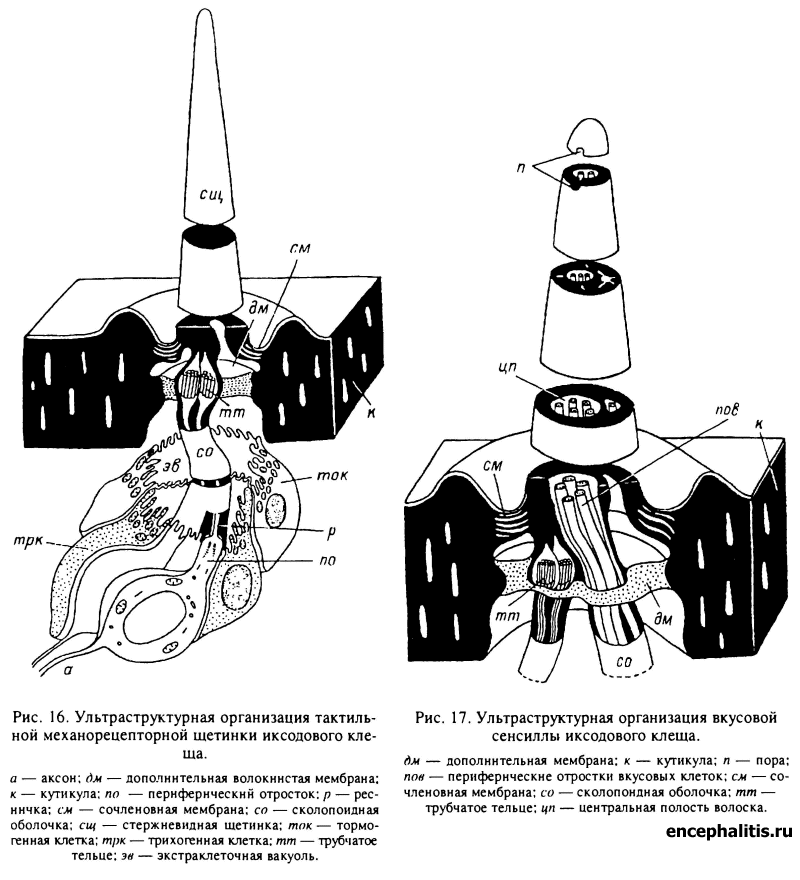
Многообразную информацию о внешней среде таежный клещ получает посредством органов чувств, среди которых ведущая роль принадлежит кутикулярным рецепторам, или сенсиллам. Все сенсиллы таежного клеща, как и у прочих членистоногих, построены по общему плану и состоят из кутикулярного отдела — тем или иным образом устроенного кутикулярного волоска (щетинки, хеты), рецепторных клеток, или биполярных чувствительных нейронов, и вспомогательных клеточных элементов (оберточных клеток), формирующих кутикулярную часть (трихогенная клетка) и сочленовную ячейку (тормогенная клетка) и обеспечивающих трофику и функционирование рецепторных нейронов (рис. 16, 17). В литературе по рецепторам кутикулярные отделы сенсилл принято называть волосками, а в литературе по систематике — щетинками, или хетами. Обладая общим планом
строения, сенсиллы разных типов (обонятельные, вкусовые, тактильные и др.) сильно различаются в деталях строения воспринимающего аппарата (щетинка, с которой связаны апикальные отделы дендритов рецепторных нейронов).
Глаза иксодовых клещей
У большинства родов Amblyomminae имеются парные глаза, расположенные латерально на границе скутума с аллоскутумом (табл. XXVI, 4). Хорошо развитыми орбитальными глазами обладают все фазы развития Н. asiaticum (Атлас..., 1979). Глазные линзы выпуклые, полусферической формы. Они представляют локальные полусферические утолщения прозрачной кутикулы, под которой видны обычные гиподермальные клетки, отделяющие ее от фоторецепторных клеток (рис. 18). Под каждой линзой располагается группа из 20—30 фоторецепторных, или зрительных, клеток, аксоны которых образуют зрительный нерв, следующий в оптический центр синганглия. Поверхность фоторецепторных клеток, противоположная стороне отхождения аксона, несет многочисленные микроворсинки. Последние представляют рабдомеры, являющиеся воспринимающими аппаратами фоторецепторных клеток. Сходным строением обладают и глаза клеща R. evertsi (Gothe et al., 1990).
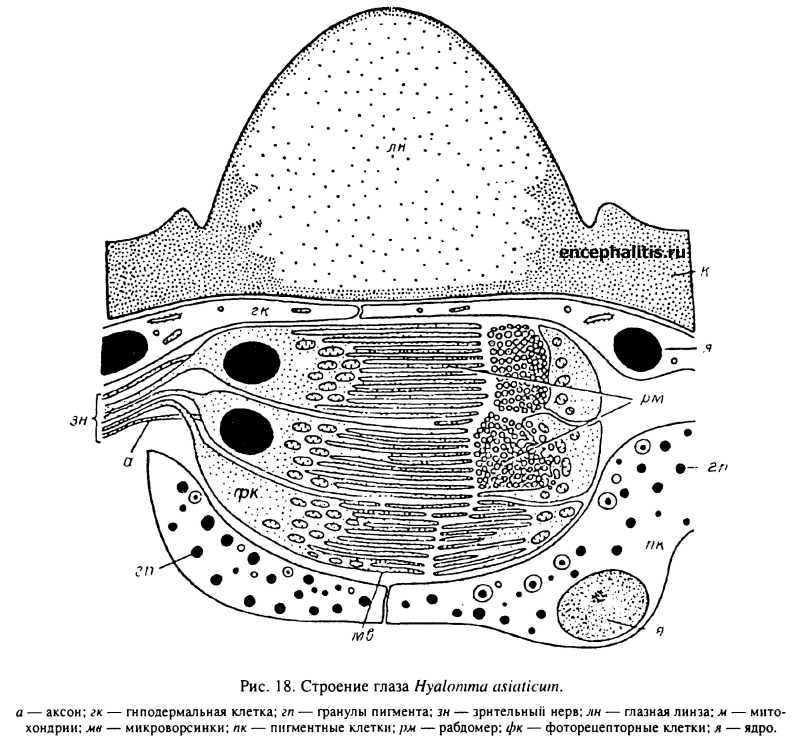
Экспериментально показано, что у клещей H. dromedarii и A, variegatum, обладающих глазами описанного выше строения, спектральная чувствительность лежит в диапазоне 340—750 нм. У первого вида обнаружены 2 максимума чувствительности при 380 и 500 нм, а у второго — только 1 при 480 нм. Для этих клещей характерна очень высокая чувствительность при низких уровнях освещенности. Оптическая ось каждого из глаз направлена в сторону и над горизонтом. Поле зрения клещей ограничено и составляет у Н. asiaticum 14° по горизонтали и 25° по вертикали, а у Amblуотта соответственно 43° и 49° (Kaltenrieder ef al., 1989).
У видов Haemaphysalis и Ixodes глаза отсутствуют, однако в поведенческих опытах с I. persulcatus отмечена реакция на свет, исчезающая при закрашивании переднего дорсального отдела идиосомы (Панфилова, 1976). У безглазых видов Ixodes, Haemaphysalis и Aponomma под кутикулой были обнаружены скопления светочувствительных клеток в тех местах, где приблизительно размещаются глаза у видов, ими обладающих (Binnington, 1972).
Тактильные сенсиллы
Идиосома, а также значительные участки поверхности гнатосомы и ног покрыты щетинками, относящимися главным образом к тактильным механорецепторным и в меньшей степени к контактным хеморецепторным сенсиллам. Тактильные щетинки (табл. II, 1, 2; IX, 1; X, 1, 2) поставляют информацию о механическом характере субстрата, например при передвижениях в подстилке; часть из них выполняет роль проприоцепторов, которые размещаются близ сочленений и сигнализируют о взаиморасположении члеников.
Принцип строения всех тактильных сенсилл сходен (рис. 16). Кутикулярная часть сенсиллы представлена жесткой стержневидной щетинкой, имеющей внутреннюю полость лишь в самой проксимальной части; иногда в эту полость заходит отросток трихогенной клетки. Сама щетинка располагается в хорошо выраженной сочленовной ячейке и присоединяется к кутикуле покровов эластичной сочленовной мембраной. В состав сенсиллы входят два биполярных рецепторных нейрона, от перикариона каждого из которых отходят центральный отросток (аксон) и периферический отросток (дендрит), идущий к основанию щетинки. Дистальный отдел периферического отростка здесь, как и во всех остальных типах сенсилл таежного клеща, представлен видоизмененной ресничкой, в базальной области имеющей строение, типичное для ресничек мерцательных эпителиев (табл. XXVII, 1, 2); апикальнее, по ходу реснички внутренняя пара бивалентов отсутствует, что весьма типично для рецепторных клеток в отличие от мерцательных (табл. XXVII, 3). Вблизи щетинки, к основанию которой ресничка жестко крепится в собственной сколопоид-ной оболочке, синтезируемой оберточными клетками, в ресничке обнаруживаются так называемые трубчатые тельца — цилиндрической формы структуры, состоящие из пучка плотно упакованных микротрубочек (табл. XXVII, 4), — типичные структуры тактильных механорецепторных нейронов членистоногих. Оберточные клетки представлены внутренней (дериватом трихогенной) и внешней (дериватом тормогенной) клетками, формирующими вокруг рецепторных ресничек при их прохождении к основанию щетинки особую экстраклеточную вакуоль.
При отклонении щетинки под действием механической силы энергия данного воздействия передается на трубчатые тельца через деформацию особой дополнительной волокнистой мембраны, связывающей эти тельца с кутикулой покровов. Обратимая деформация мембран трубчатых телец служит пусковым механизмом возбуждения механорецепторного нейрона.
К чувствительным образованиям у клещей относят тарзальные железы. Однако роль рецепторных элементов, анатомически входящих в состав сложных желез, которые обслуживают подушечки конечностей клеща, остается неясной.
Контактные хеморецепторы, или органы вкуса, обнаружены на лапках, пальпах, хелицерах, основании гнатосомы; входят в состав пальпального рецепторного органа, подробно исследованного у клеща Н. asiaticum (Атлас..., 1979).
Кутикулярный отдел сенсилл данного типа представлен полым толстостенным волоском базиконического типа, длина которого варьирует в зависимости от месторасположения на теле клеща. Полый базиконический волосок вблизи вершины перфорирован одной или несколькими относительно крупными порами; иногда его стенки пронизаны тонкими извитыми канальцами (рис. 17; табл. XXVIII, 1). К подобной щетинке через канал в кутикуле покровов подходят две группы рецепторных ресничек, следующих каждая в собственной сколопоидной оболочке. Реснички, идущие от группы, которая состоит из двух клеток, жестко крепятся у основания волоска, образуя трубчатые тельца, следовательно, по морфологическим признакам относятся к механорецепторам (рис. 16). «Чистые», т. е. лишенные механорецепторов хеморецепторные сенсиллы у клещей, как и у других членистоногих, неизвестны. Реснички периферических отростков второй группы, в состав которой может входить от 1—2 до 5 клеток в разных сенсиллах, проходят во внутренний канал полого базиконического волоска и следуют, не ветвясь, до района расположения пор. Возможность химической стимуляции рецептивных участков мембран ресничек через поры обеспечивается лишь при контакте вершины щетинки с субстратом.
Коготковые сенсиллы
В основании коготка I пары ног амблиомин располагаются симметрично 3 пары волосков, называемых коготковыми сенсиллами, а у иксодин — 2 пары. По своему строению они относятся к хеморецепторным контактным сенсиллам и сходны с подобными волосками пальпального органа. Предполагают, что у самцов клещей они служат рецепторами мало летучих половых феромонов самок (Philips, Sonenshine, 1993).
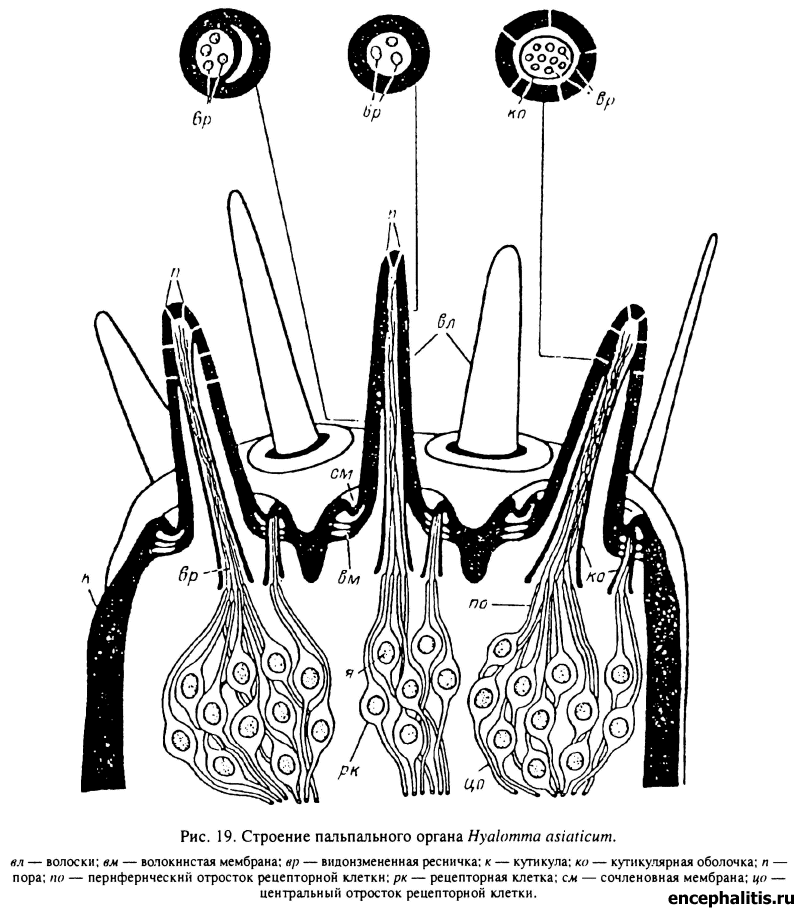
Пальпальный орган, как принято называть скопление чувствительных щетинок (базиконических волосков) на вершине IV членика пальп (рис. 19; табл. XXIX, 2; XXX, 1), у половозрелых клещей образован 10 хеморецепторными сенсиллами контактного типа. Группа базиконических волосков на вершине пальпы окружена 9 более тонкими щетинками — предположительно контактными механорецепторами. Базиконические волоски пальпального органа морфологически подразделяются на два типа, различающихся строением пор и канальцев в кутикуле, что позволяет предполагать функциональную специализацию сенсилл пальпального органа (Балашов и др., 1976).
Хелицеральные сенсиллы
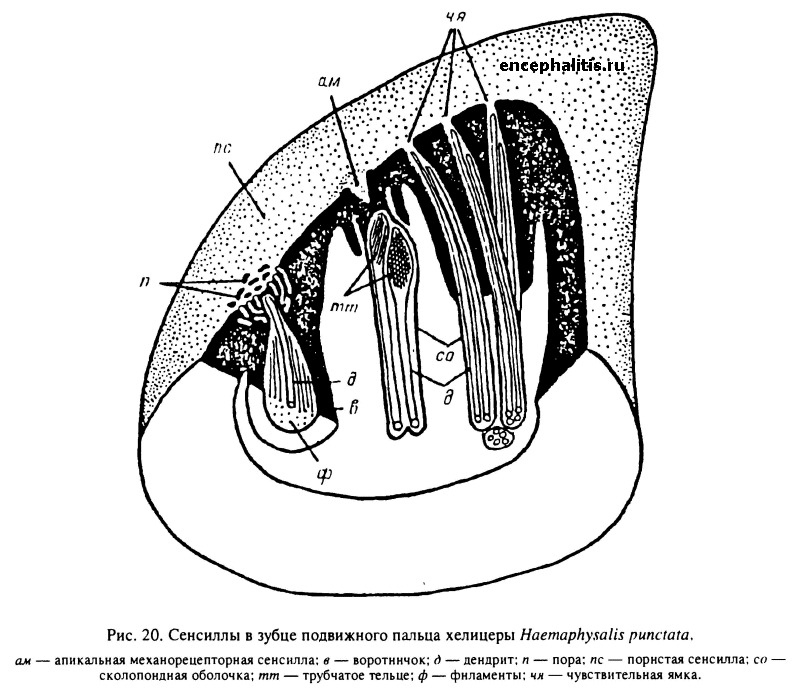
Большое количество сенсилл обнаружено на пальцах хелицер (Waladde, Rice, 1982; Данилов, 1987а, 19876, 1988). У видов родов Boophilus, Dermacentor, Haemaphysalis, Hyalomma выявлено 11 сенсилл с 29—30 чувствительными клетками в каждой из хелицер, а у I. persulcatus и I. uriae — 12 сенсилл с 34 клетками. Число сенсилл и связанных с ними чувствительных клеток одинаково у самцов и самок одного вида. У всех изученных видов в зубце подвижного пальца обнаружено 5 сенсилл (рис. 20; табл. XXIX, 4). Среди них различают апикальную механорецепторную сенсиллу, хеморецепторные сенсиллы (чувствительные ямки) и пористую сенсиллу, служащую терморецептором. В основании подвижного пальца с медиальной стороны расположена медиальная механорецепторная сенсилла, а с латеральной стороны — латеральная механорецепторная сенсилла. На неподвижном пальце находятся 4 или 5 хеморецепторных сенсилл, называемых чувствительными ямками.
Функциональное значение контактных хеморецепторов клещей состоит главным образом в поиске места для кровососания на хозяине и в стимуляции самого этого процесса. Прицепившись к прокормителю, клещ при помощи вкусовых сенсилл на лапках и пальпах определяет места непосредственного контакта с кожными покровами млекопитающего. После этого по сочетанию ряда признаков (выделению потовых и иных желез и др.) и, вероятно, при участии терморецепторов он определяет оптимальное место для присасывания. Непосредственно с внутренними слоями кожи, кровью и лимфой контактируют органы вкуса, расположенные на хелицерах. Они, по-видимому, стимулируют начало кровососания после прорезания кожи и могут определять состав поглощаемой пищи.
Орган Галлера
Главным дистантным рецепторным органом, определяющим приближение к клещу потенциального прокормителя, служит полифункциональный (преимущественно обонятельный) орган Галлера, размещающийся на дистальной поверхности передних лапок клеща (табл. XXVI, 1, 2; XXX, 2). Строение этого исключительно сложного образования достаточно подробно описано в литературе (Леонович, 1977, 1978; Балашов, Леонович, 1978, 1981; Атлас..., 1979).
Основным отделом органа Галлера является обонятельная капсула, которая представляет собой углубление в кутикуле дорсальной поверхности I пары лапок, закрытое сверху крышкой с небольшим отверстием (рис. 21). Внутри капсулы размещены пористые обонятельные волоски. У видов Amblyomminae кроме волосков в капсуле находятся и кутикулярные выросты неправильной формы, называемые плеоморфами, и могут открываться протоки нескольких двухклеточных желез.
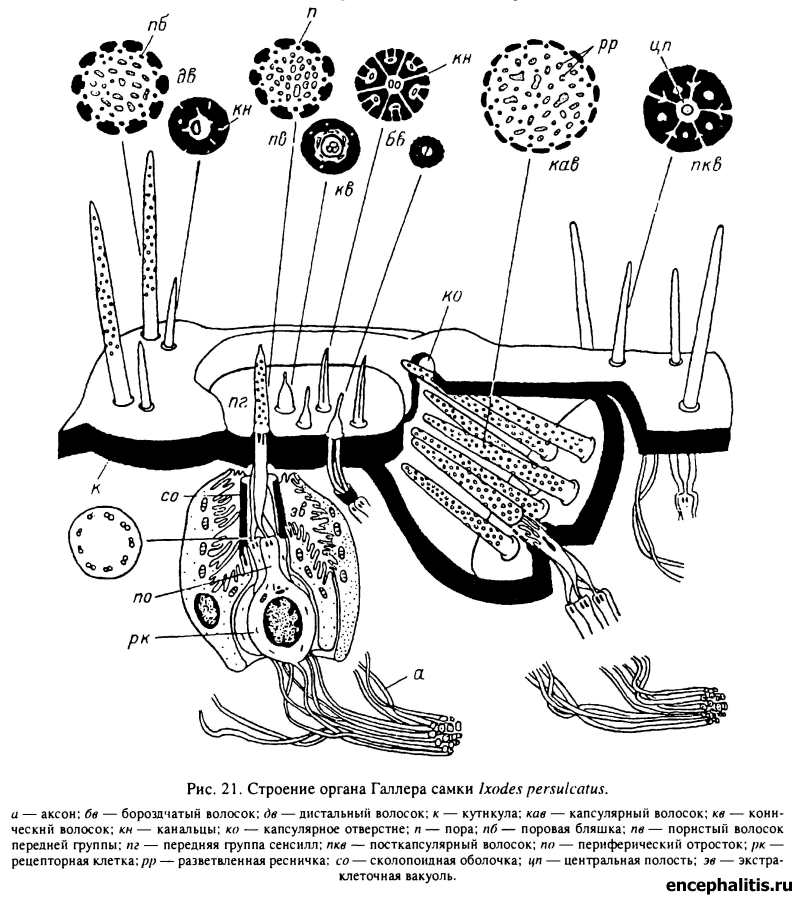
Каждая сенсилла капсулы включает по 4—5 биполярных чувствительных нейронов и 2 оберточные клетки. Главная особенность воспринимающего аппарата капсулярных сенсилл — наличие полого тонкостенного кутикулярного волоска, стенки которого пронизаны многочисленными порами, а также обильные ветвления ресничек в полости волоска.
Дистальнее капсулярного отверстия в обширном, хорошо выраженном углублении размещен второй отдел органа Галлера — передняя группа сенсилл (табл. XXVI, 3; XXX, 3). У I. persulcatus она представлена 6 волосками 4 типов: одним пористым, двумя бороздчатыми, двумя тонкими и одним коническим волоском.
Пористый волосок, иннервируемый 6 рецепторными клетками, сходен строением с обонятельными волосками капсулы, но отличается от них большей толщиной стенок (табл. XXVIII, 2; XXX, 4). Два волоска такого же строения входят в состав группы так называемых дистальных сенсилл, размещающихся дистальнее передней группы сенсилл.
Весьма своеобразно устроены бороздчатые волоски, несущие на поверхностях продольные бороздки (насечки) (табл. XXVI, 3; XXX, 4). Их воспринимающий отдел представлен кутикулярным волоском со сложной системой полостей и канальцев: в центральной полости проходят, не ветвясь, рецепторные реснички 2—3 чувствительных нейронов, а в окружающих концентрически центральную полость периферических полостях проходят отростки оберточных клеток. Как центральные, так и периферические полости сообщаются со средой посредством тонких канальцев (табл. XXVIII, 3). Конические и два тонких волоска устроены проще.
Непосредственно за капсулярным отверстием размещается группа из нескольких «их число может варьировать) посткапсулярных волосков (табл. XXX, 2), каждый из которых иннервируется 1—2 клетками. Реснички периферических отростков этих клеток проходят в центральной полости волоска, соединяющейся со средой канальцами и системой щелевидных полостей. Концентрические периферические полости сообщения со средой не имеют (табл. XXVIII, 4). По обеим сторонам органа Галлера обнаруживаются вкусовые сенсиллы обычного строения.
Различия в строении волосков передней группы сенсилл, посткапсулярных волосков и волосков капсулы предполагают различие в их функциях. Обонятельная функция капсулы органа Галлера в восприятии летучих компонентов «запаха хозяина» была впервые показана для I. ricinus в поведенческих опытах (Lees, 1948), и в дальнейшем многообразные функции этого органа были выявлены специальными электрофизиологическими и поведенческими исследованиями (Guerin et al., 1992). В частности, для A. variegatum установлено, что среди тонкостенных пористых волосков капсулы рецепторные клетки одного возбуждаются, а другого ингибируются определенными концентрациями углекислого газа в воздухе (Steullet, Guerin, 1992a). Еще 2 волоска являются рецепторами сульфидов и аммиака (Steullet, Guerin, 1992b). Среди чувствительных клеток волосков капсулы также выявлены рецепторы бензальдегидов, алифатических альдегидов и лактонов, выделяемых при дыхании и испускаемых поверхностью тела (Steullet, Guerin, 1994a).
Среди волосков передней группы органа Галлера один наиболее крупный воспринимает нитрофенол — главный компонент половых феромонов клещей. Кроме того, в этой группе сенсилл обнаружены рецепторы различных компонентов запаха хозяина (лактонов, жирных кислот, аммиака, метилпропановой и бутановой кислот) (Steullet, Guerin, 1994b).
Определенное сходство в ультраструктуре бороздчатых и посткапсулярных волосков с таковой термогигрорецепторов ряда членистоногих заставляет предположить, что функцией таких волосков может быть термо- и гигрорецепция (Леонович, 1977). Реакция таежного клеща на температуру (инфракрасное излучение) показана в полевых экспериментах (Елизаров, Васюта, 1976), а обитание и преимущественное тяготение этого вида к мезофильным биотопам свидетельствуют о наличии рецепторов, сигнализирующих об уровне влажности.