Иксодовые клещи — паразиты и переносчики инфекций
Глава II. РАСПРОСТРАНЕНИЕ
Иксодовые клещи характеризуются всесветным распространением, а один из видов (I. uriae) встречается даже на островах и побережьях Арктики и Антарктики. Наибольшим видовым разнообразием отличается фауна иксодид тропических и субтропических регионов. Географическое распространение иксодид как временных эктопаразитов зависит от условий окружающей среды, распространения их прокормителей и отражает историю формирования фауны конкретного региона.
Проводя большую часть своей жизни вне тела хозяина, иксодовые клещи, как и другие свободноживущие организмы, зависят от совместного воздействия множества абиотических и биотических факторов. Географические ареалы видов этих клещей во многом соответствуют распределению в пространстве условий среды, пригодных для их существования. При отсутствии географических преград распространение некоторых видов в значительной степени обусловлено гигротермическими условиями.
Сходная картина зависимости распространения иксодид от климатических факторов обнаружена для нескольких видов, и на этом основании многие исследователи полагают, что пространственное расселение иксодид в целом совпадает с соответствующими климатическими, ландшафтными, геоботаническими и другими характеристиками территорий. Эти постулаты были положены в основу создания карт распространения отдельных видов клещей на основе уже существующих природно-климатических карт разного масштаба, часто при очень ограниченных данных о реальных находках этих животных в природе. Подобные карты дают приблизительные контуры видовых ареалов, но не могут воспроизвести реальное расселение вида на занимаемой им территории и особенно свойственный многим иксодидам мозаичный или пятнистый тип расселения.
К настоящему времени с разной степенью точности установлено географическое распространение большинства видов иксодид, и эти материалы достаточно полно собраны Г. В. Колониным (1978, 1981, 1983, 1984). Однако причины различий в фаунах клещей зоогеографических регионов невозможно понять без анализа эволюции иксодид на уровне родов и подродов и путей расселения этих таксонов в прошлом (Балашов, 1993а). При использовании зоогеографических данных следует допустить ограниченную способность этих клещей к преодолению морских и океанических пространств, разделяющих континенты, и в меньшей степени гор и пустынь. Даже в случае межконтинентальных заносов иксодид с мигрирующими птицами, регулярно происходящих между Европой, Африкой и Азией, мигранты обычно неспособны к закреплению в необычных для них геобиоценозах. Для ареалов иксодид характерно постепенное расширение или сужение границ в ответ на соответствующие многолетние изменения биотических и абиотических факторов, а не взрывная экспансия.
Исторические факторы в расселении иксодид отражают сложные процессы развития отдельных таксонов в условиях изоляции континентов и обмена фаун при соединениях континентальных платформ. В то же время на обширных массивах суши, как например Евразия или Северная и Южная Америка, распространение клещей определяется наряду с таковым их прокормителей абиотическими факторами среды и в первую очередь температурой и влажностью (табл. 1).
Наиболее морфологически примитивными и древними по происхождению среди Ixodidae считают в подсем. Ixodinae единственный род Ixodes, а в подсем. Аmblyomminae — роды Аропотта и Amblyomma. Эти 3 рода имеют космополитические ареалы, причем виды Ixodes распространились во всех климатических поясах, а два других рода — только в тропиках и субтропиках. По-видимому, предки этих родов существовали еще до распада единого массива суши — Пангеи на северную половину — Лавразию и южную — Гондвану. Это событие датируется серединой юры (Ушаков, Ясаманов, 1984; Оуэн, 1988), так что роды Ixodes, Аропотта и Amblyomma должны были существовать с середины мезозоя.
Дальнейшая судьба этих групп оказалась неодинаковой. Виды рода Аропотта (23) обитают преимущественно в разных типах тропических и субтропических лесов Восточного полушария (21 вид). Только в Новой Зеландии A. sphenodonti — паразит гаттерий — живет в умеренном климате, а в Австралии A. hydrosauri приспособился к аридным условиям пустынь. Наиболее богата фауна Азии (12 видов), в Австралии обитает 6 видов, в Африке — 4 и в Центральной и Южной Америке — 2 (рис. 26) (Колонии, 1983).
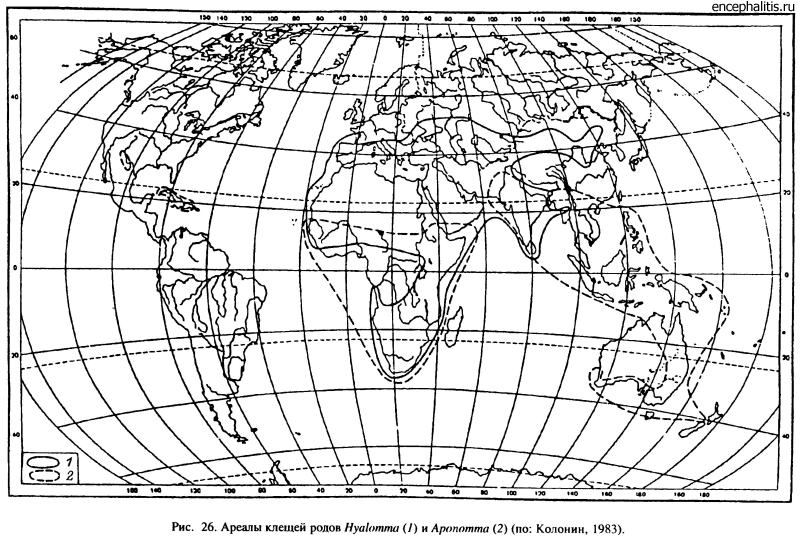
Род Amblyomma, включающий 102 вида, оказался более пластичным как в выборе прокормителей (рептилии, млекопитающие, птицы), так и по отношению к абиотическим факторам. Местообитаниями значительной части видов служат влажные тропические и субтропические леса, но большая часть африканских видов (A. variegatum, A. hebraeum, A. tholloni и др.) приспособилась к существованию в саваннах (Petney et al., 1987). В Южной и Центральной Америке также есть ксерофильные виды, встречающиеся в прериях (A. parvum, A. usingeri и др.). В зону с умеренным климатом заходят только A. americanum до 40° с. ш. в США (рис. 27) и A. parvitarsum, достигающий в Патагонии 50° ю. ш. Все остальные виды ограничены в своем распространении областями с тропическим и субтропическим климатом.
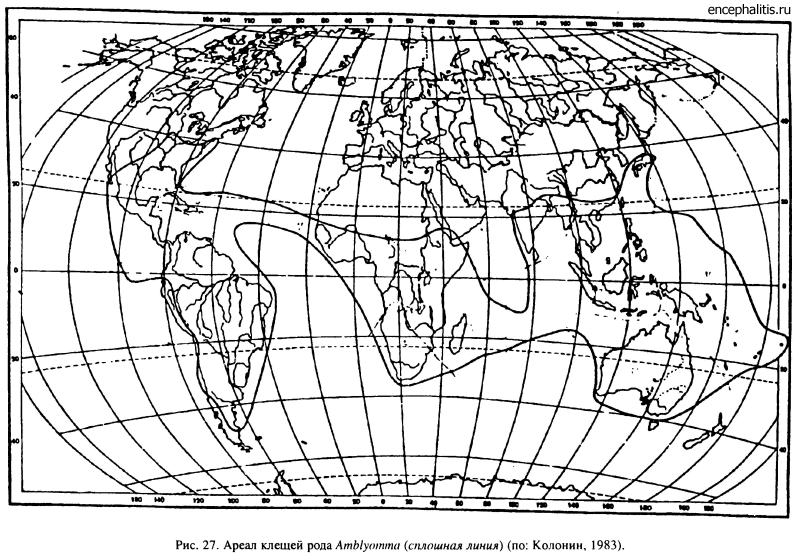
Наибольшее число видов (52) свойственно фауне Южной и Центральной Америки, включая Антильские острова и острова Галапагос с многочисленными эндемиками. Общие виды для Восточного и Западного полушарий отсутствуют, за исключением недавно завезенного из Африки на Антильские острова A. variegatum. В Африке обнаружено 19 видов, включая один эндемичный для Мадагаскара. В странах Южной и Юго-Восточной Азии встречается 17 видов и в Австралии — 12 (Колонии, 1983).
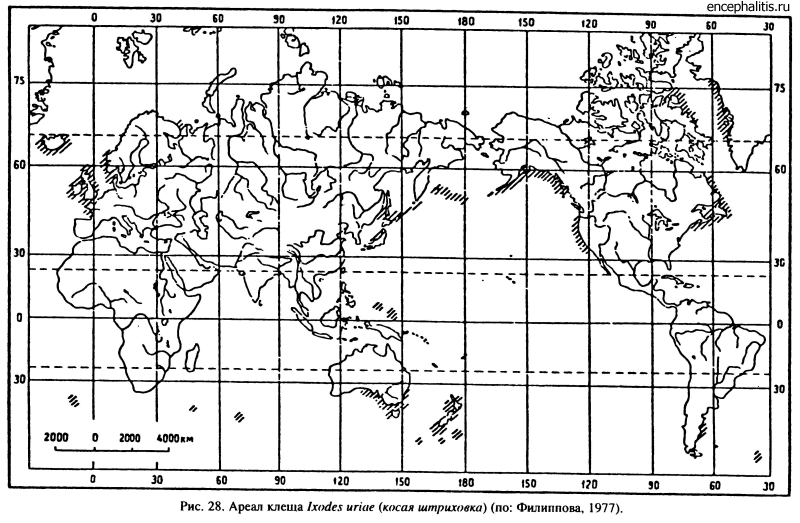
Самый многочисленный в семействе род Ixodes (около 250 видов) встречается во всех зонах вплоть до побережий полярных морей. Согласно Н. А. Филипповой (1977), внутри рода, включающего 19 подродов, имеются как более примитивные, так и эволюционно продвинутые группы. К числу первых принадлежат подроды Ceratixodes, Scaphixodes, Multidentatus, паразитирующие на древнейших по происхождению морских птицах, остатки которых известны с палеогена. Всесветные ареалы хозяев обусловили подобное же распространение их паразитов, не помогающее филогенетическим реконструкциям (рис. 28).
Из паразитов наземных млекопитающих наиболее морфологически примитивны подроды Sternalixodes, Exopalpiger, Endopalpiger, большая часть видов которых встречается только в Австралии, но по 1 виду подрода Exopalpiger встречается в Южной Америке, Африке и в Евразии. Судя по рецентным ареалам видов этого реликтового подрода, он должен был возникнуть в юре до распада Пангеи, и до настоящего времени эти клещи сохранили трофические связи с наиболее примитивными группами млекопитающих — однопроходными, сумчатыми и насекомоядными.
В пользу большой древности рода Ixodes свидетельствует и распространение наиболее богатого видами и эволюционно продвинутого подрода Ixodes s. str., отсутствующего только в Африке к югу от Сахары и в Австралии. По-видимому, этот подрод сформировался уже после распада Пангеи в ее северной половине — Лавразии до утраты наземного соединения Северной Европы, Америки и Европы, т. е. не позднее конца палеоцена. Дальнейшее расселение этих клещей шло в южном направлении на соединившийся с Азией в конце олигоцена — начале эоцена Индийский субконтинент, позднее в миоцене через Иберийский мост в Северную Африку, а из Северной Америки в Южную в плиоцене через образовавшийся тогда Панамский перешеек или же раньше, через острова Карибского моря. Последний путь менее вероятен, так как виды этого подрода не смогли преодолеть морские проливы между Южной Азией и Австралией.
Несмотря на широкое распространение, виды рода Ixodes повсеместно приурочены к местообитаниям с достаточной влагообеспеченностью. Виды, подстерегающие прокормителей на поверхности почвы или растительности, как например I. persulcatus и I. ricinus, приурочены к различным типам леса, где постоянно поддерживается высокая относительная влажность в приземном слое воздуха. В степях, саваннах и пустынях, где относительная влажность воздуха испытывает значительные сезонные и суточные колебания и может быть очень низкой, встречаются только виды с гнездово-норовым паразитизмом, который, по-видимому, возник независимо в нескольких подродах (Ixodes s. str., Afrixodes и др.). Примером подобных видов в Палеарктике служат I. laguri, I. subterraneus или I. осculms. Некоторые гнездово-норовые виды приспособились к жизни в околоводных норах с избыточным увлажнением, как например I. apronophorus в Палеарктике, I. ornithorynchi из нор утконосов в Австралии или I. aulacodi, паразитирующий в норах тростниковой крысы в Африке. Подобные же условия избыточной увлажненности характерны для местообитаний паразитов колониальных морских птиц — I. uriae, I. signatus и др.
Широкое распространение гнездово-норового паразитизма в роде Ixodes, вероятно, вторично, хотя и очень древнего происхождения. Ему должна была предшествовать стадия пастбищного паразитизма, обеспечившая выработку «жизненной формы» иксодового клеща как кровососа с чередованием стадий многодневного питания и еще более продолжительного свободного существования. На этой стадии эволюции должен был выработаться весь комплекс морфофизиологических и экологических адаптации, большинство из которых сохраняется при последующем переходе к гнездово-норовому паразитизму. Последний, несомненно, возникал неоднократно внутри разных эволюционных линий и встречается как в примитивных подродах (Coxixodes, Ceratixodes, Pholeoixodes и др.), так и в наиболее эволюционно продвинутом подроде Ixodes s. str. (I. apronophorus, I. laguri, I. occultus), в котором доминирует пастбищный тип паразитизма. В связи с этим следует отметить, что значительная часть видов морфологически примитивных подходов Exopalpige'r, Endopalpiger, Sternalixodes представлена не типичными гнездо-ю-норовыми, а пастбищными паразитами, как например австралийский I. (St.) holocyclus. У относящегося к этой группе I. (Ex.) trianguliceps, в частности, выявлен «примитивный» тип пастбищного паразитизма, при котором активные перемещения клещей ограничены самым верхним слоем почвы, растительной подстилкой и припочвенным ярусом растительности.
Второй по численности в семействе род Haemaphysalis (147 видов), согласно взглядам Хугстрала (Hoogstraal, Kim, 1985), включает 16 подродов, из которых 4 из 17 видов считаются эволюционно примитивными, а 12 со 130 видами — эволюционно продвинутыми. Подавляющая масса видов (более 90) обитает в Индо-Малайской области. В Эфиопской области встречается 19 видов и еще 12 эндемичных видов на Мадагаскаре, в Австралии — 12 видов, в Палеарктике — 19 и в Северной и Южной Америке — 4.
Большинство видов приурочено к разным типам зональных лесов тропической и субтропической зон. В умеренном климате при общем обеднении фауны эти клещи также встречаются в разных типах лесных местообитаний, причем могут иметь обширнейшие ареалы. Например, Н. concinna распространен от Западной Европы до Японии (рис. 33), а Н. leporispalustris от Аляски до Аргентины. Многие виды обитают в горных лесах. Общей особенностью всех этих местообитаний является достаточно высокая и стабильная влажность. Лишь немногие виды приспособились к существованию в аридных местообитаниях, как например H. рипctata, встречающийся в редколесьях и кустарниковых зарослях, или H. sulcata, встречающийся в степных и полупустынных горных районах Южной Европы, Передней и Средней Азии. Такие виды, как Н. erinacei или Н. hispanica из подрода Rhipistoma, обитают в ксерофильных ландшафтах, перейдя к гнездово-норовому паразитизму. На связь клещей рода Haemaphysalis с мезофильными лесными ландшафтами было впервые указано Б. И. Померанцевым (1948а), но вопрос о месте и времени происхождения и дальнейшей эволюции рода остается дискуссионным.
Хугстрал (Hoogstraal, Kim, 1985) предполагает, что род Haemaphysalis возник на Индостанском субконтиненте, откуда клещи в дальнейшем распространились в северо-западном направлении, достигнув побережий Атлантического и Тихого океанов. Они расселились по Африке, но лишь немногие смогли проникнуть в Северную и Южную Америку.
По нашему мнению (Балашов, 1993а), исходя из современных мобилистических представлений, подобная трактовка происхождения ареалов является спорной. Первоначально Индия была частью Гондваны, в мелу отделилась от нее и на протяжении многих миллионов лет оставалась изолированным островом. Столкновение Индии с Евразией произошло в конце олигоцена или начале эоцена и привело к мощным процессам горообразования, завершившимся образованием Гималаев. Гондванские корни рода представляются маловероятными, так как из других частей Гондваны Южная Америка, несмотря на обширную область тропических лесов, наиболее бедна представителями этого рода. Сравнительно небогаты фауны Haemaphysalis Австралии и Африки, причем на все 3 континента клещи несомненно проникли с севера. В связи со сказанным мы полагаем, что наиболее вероятны происхождение и ранние этапы эволюции рода в палеогене на территории Юго-Восточной Азии, где тропический климат и влажные леса сохранялись на протяжении всего кайнозоя. В пользу этой гипотезы свидетельствует распространение наиболее примитивных 3 из 4 современных видов подрода Allocerae в горных субтропических лесах этого региона, и только 1 — Н. inermis в лесах Южной Европы и Передней Азии. Первоначально эти клещи могли проникнуть в умеренно теплые части Евразии. Распространение клещей в Индостан и Африку произошло позднее, после их соединения с Евразией, а в Австралию они были занесены из Юго-Восточной Азии с птицами или, возможно, с грызунами сем. Muridae, сумевшими преодолеть морские проливы.
Из районов с тропическим и субтропическим климатом наиболее раннее вселение клещей произошло в Индию, и поэтому она богата видами этого рода по сравнению с другими осколками Гондваны. Крайняя бедность фауны Южной Америки может быть объяснена отсутствием клещей в фауне Северной Америки, куда они могли проникнуть через сравнительно холодную Берингийскую сушу и затем, двигаясь на юг через Панамский перешеек, попасть в Южную Америку. Эти события могли произойти только в плиоцене всего несколько миллионов лет назад, так что времени на образование эндемичной фауны оказалось недостаточно.
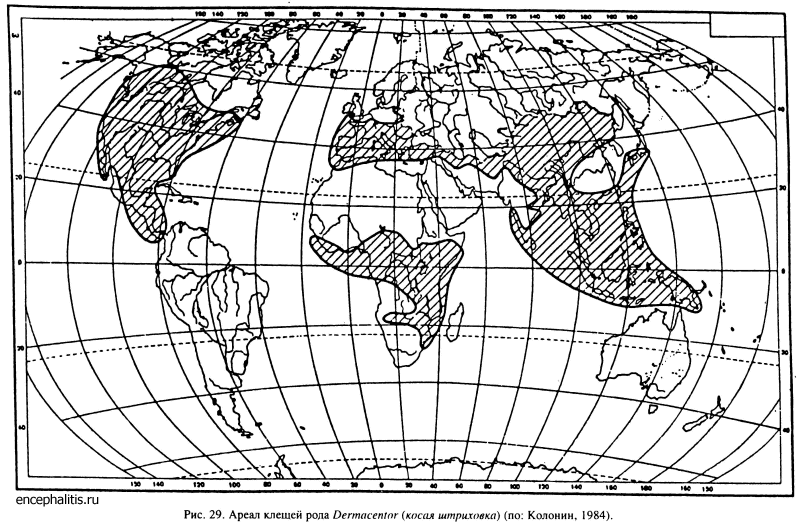
Очень обширным ареалом обладает род Dermacentor, насчитывающий около 32 видов (рис. 29). В Палеарктике обитает 15 видов, в Северной и Центральной Америке вплоть до 10° ю. ш. — 11 видов, в Тропической Азии — 4 вида и в Африке к югу от Сахары — 2 вида. Померанцев (19486) полагал, что этот род возник в горах или степях Центральной Азии и позднее по Берингийской суше проник в Северную и далее в Центральную Америку. Распространение этого вида в тропические области Старого Света происходило сперва с востока в Ориентальную область, а из нее в Эфиопскую. Согласно другой точке зрения (Белозеров, 1976; Колонии, 1984), палеарктические представители рода имеют североамериканское происхождение, а Евразийский центр видообразования этой группы вторичен.
Обе рассматриваемые гипотезы обходят стороной вопрос о хронологии описываемых процессов, но сходятся на единственно возможном пути расселения через Берингийскую сушу. Последняя возникала и исчезала неоднократно на протяжении кайнозоя, причем последние «мосты» датируются плиоценом-плейстоценом с возрастом в 3.5, 1.8 и 0.08 млн лет назад. Наиболее ранние мосты были возможны в конце мела или в плиоцене около 60 млн лет назад. Современные представления мобилистической геологии позволяют дать и третье возможное объяснение расселению рода Dermacentor, во многом сходное с таковым у хвостатых амфибий. Мы полагаем, что преимущественное распространение этих клещей в умеренных и умеренно теплых климатических зонах северного полушария и отсутствие в таковых южного допускают возникновение этих клещей в Лавразии уже после ее разделения с Гондваной, т. е. не ранее мела. В дальнейшем представители рода могли успеть расселиться по Евразии и Северной Америке еще до их расхождения и утраты соединения через Гренландию в палеоцене 45—50 млн лет назад, так что изоляция фаун этих континентов очень древняя. Широкому распространению клещей в южном направлении, скорее всего, препятствовали экологические факторы, и немногие виды, приспособившиеся к существованию в тропиках Юго-Восточной Азии и Африки, выделены систематиками в подроды Indocentor и Amblуocentor. Крайняя бедность фауны Южной Америки легко объяснима ее островным положением до плиоцена, так что немногие виды смогли проникнуть в этот регион не ранее 3 млн лет назад при массовом вселении в нее североамериканских млекопитающих через вновь образовавшийся Панамский перешеек. Возможно, что только D. nitens, иногда выделяемый в самостоятельный род Anocentor и широко распространенный в тропических и субтропических районах, мог проникнуть в Неотропическую область значительно раньше через острова Карибского моря вместе с ранними млекопитающими-вселенцами. Подобный путь распространения, в частности, допускается Траубом (Traub, 1980) для объяснения особенностей фаун блох и вшей этого региона.
Для палеарктических и большей части голарктических видов характерна приуроченность к различного рода степным или луговым ландшафтам, причем заметна связь многих из них с горным ландшафтом, а такие виды, как D. pavlovskyi или D. montanus, поднимаются в горах Средней Азии до верхних границ растительности на высотах 3500—4000 м над ур. м. (Филиппова, Панова, 1989). Встречающиеся в лесных ландшафтах клещи D. reticulatus и D. silvarum заселяют разного рода редколесья, вырубки, опушки и другие интразональные элементы. D. niveus заселил поймы рек с тугайной растительностью в аридной зоне. Можно предполагать, что связи этих клещей со степными и горно-степными ландшафтами, как предполагал Померанцев (19486), первичны, а с тропическими лесами — вторичны.
Из 73 видов рода Rhipicephalus не менее 55 встречается только в Африке, 8 — в Палеарктике и 4 — в Тропической Азии. В Австралии, Южной и Северной Америке, за исключением недавно завезенного R. sanguineus, клещи этого рода отсутствуют (рис. 30). Подавляющая масса видов заселяет разные типы степей, саванн и реже пустынь. В этих стациях клещи могут заселять интразональные редколесья и кустарниковые заросли, как например обитающий в тугаях R. leporis. Несколько видов обитает во влажных тропических лесах (R. cliffordi, R. complanatus, R. ziemanni). Гнездово-норовый паразитизм свойствен немногим видам (R. schulzei и R. pusillus).
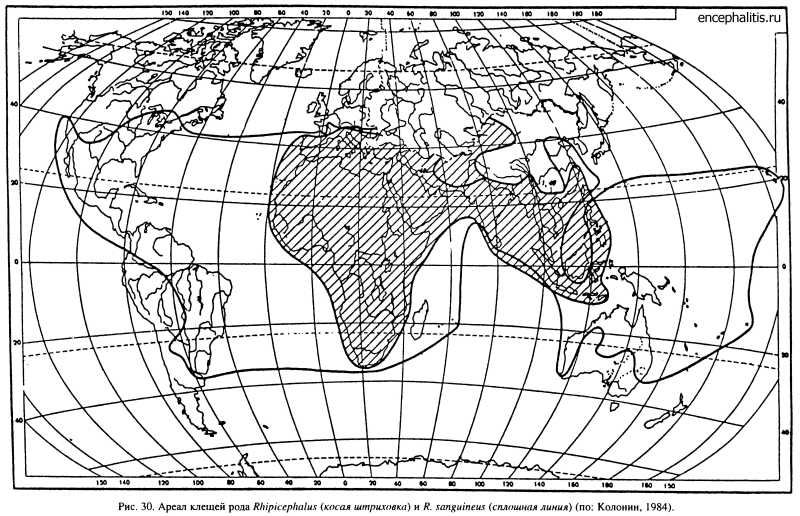
Происхождение этого рода связывают с Африкой (Померанцев, 19486; Pegram, 1984), и эта гипотеза подтверждается геологическими данными. Известно, что с олигоцена Африка вместе с Аравийским полуостровом примерно 25 млн лет оставалась островом. Только 17 млн лет назад северо-восточный угол континента соединился через Аравию с турецкой и иранской частями Евразии, а 12—14 млн лет назад произошло соединение Африки с Иберийским полуостровом. Можно предполагать, что род Rhipicephalus сформировался в период островного существования Африки, а позднее, после соединения ее с Евразией, вместе с копытными и другими аборигенами африканской фауны распространился в аридные области Палеарктики. Там, в пустынях и степях Передней и Средней Азии, согласно Померанцеву (1948б), возник более молодой вторичный очаг видообразования.
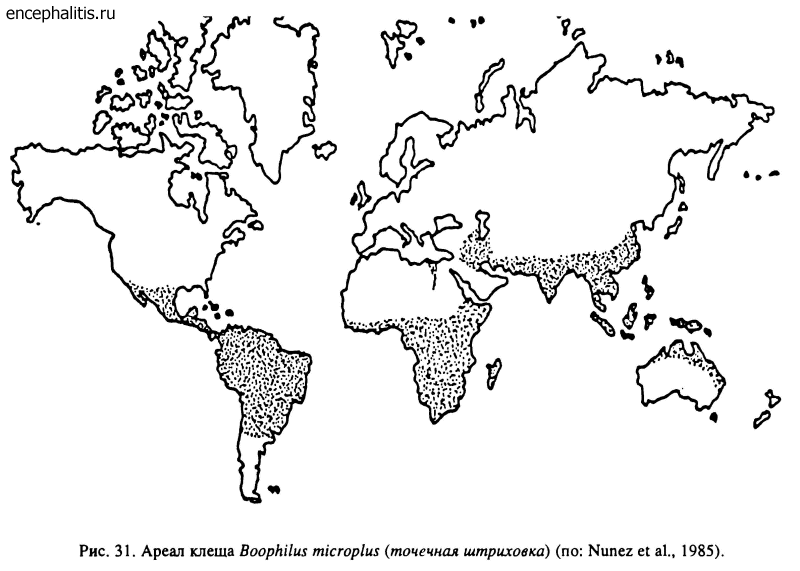
Род Boophilus включает всего 5 видов, которые до завоза с домашним скотом в последние 200 лет отсутствовали в Австралии, Северной и Южной Америке. Предполагают (Hoogstraal, 1978), что первоначально распространение В. microplus было ограничено тропическими и субтропическими районами Азии, а в историческое время он проник в Америку, Африку и Австралию (рис. 31). В. kohlsi и В. annulatus происходят из Средиземноморья, а В. geigy и В. decoloratus — из Африки к югу от Сахары.
Главными прокормителями этих клещей повсеместно служат полорогие, и можно с достаточным основанием считать это семейство парнокопытных первичными хозяевами рода. Согласно палеонтологическим данным (Thenius, 1980), полорогие появились в Европе в раннем миоцене, но адаптивная радиация и расселение этого семейства, включая проникновение в Африку и Южную Азию, происходили в плиоцене. По-видимому, возраст рода, отличающегося высокой хозяинной специфичностью, не может быть старше, чем таковой его прокормителей.
Род Hyalomma включает около 30 видов, из которых примерно половина встречается в аридных местообитаниях Северной Африки, Южной Европы, Передней и Средней Азии (рис. 26). В пустыни Центральной Азии проникают только 3 вида этого рода. В саваннах и тропических редколесьях Африки к югу от Сахары обитает 5 видов и в сходных ландшафтах Индии — 5. Померанцев (19486) связывает происхождение рода Hyalomma с пустынями Передней Азии в эоцене, откуда эти клещи распространились в более мезофильные ландшафты Индии и Африки к югу от Сахары. Подобная точка зрения представляется более правдоподобной, чем взгляды Хугстрала (Hoogstraal, Kim, 1985) на возникновение этой группы еще в мелу. Независимо от мелового или более позднего происхождения их проникновение в Индию и Африку могло произойти не раньше неогена после соединения этих массивов суши с Евразией.
Распространение иксодовых клещей
Иксодовые клещи характеризуются всесветным распространением, а один из видов (I. uriae) встречается даже на островах и побережьях Арктики и Антарктики. Наибольшим видовым разнообразием отличается фауна иксодид тропических и субтропических регионов. Географическое распространение иксодид как временных эктопаразитов зависит от условий окружающей среды, распространения их прокормителей и отражает историю формирования фауны конкретного региона.
Проводя большую часть своей жизни вне тела хозяина, иксодовые клещи, как и другие свободноживущие организмы, зависят от совместного воздействия множества абиотических и биотических факторов. Географические ареалы видов этих клещей во многом соответствуют распределению в пространстве условий среды, пригодных для их существования. При отсутствии географических преград распространение некоторых видов в значительной степени обусловлено гигротермическими условиями.
Сходная картина зависимости распространения иксодид от климатических факторов обнаружена для нескольких видов, и на этом основании многие исследователи полагают, что пространственное расселение иксодид в целом совпадает с соответствующими климатическими, ландшафтными, геоботаническими и другими характеристиками территорий. Эти постулаты были положены в основу создания карт распространения отдельных видов клещей на основе уже существующих природно-климатических карт разного масштаба, часто при очень ограниченных данных о реальных находках этих животных в природе. Подобные карты дают приблизительные контуры видовых ареалов, но не могут воспроизвести реальное расселение вида на занимаемой им территории и особенно свойственный многим иксодидам мозаичный или пятнистый тип расселения.
К настоящему времени с разной степенью точности установлено географическое распространение большинства видов иксодид, и эти материалы достаточно полно собраны Г. В. Колониным (1978, 1981, 1983, 1984). Однако причины различий в фаунах клещей зоогеографических регионов невозможно понять без анализа эволюции иксодид на уровне родов и подродов и путей расселения этих таксонов в прошлом (Балашов, 1993а). При использовании зоогеографических данных следует допустить ограниченную способность этих клещей к преодолению морских и океанических пространств, разделяющих континенты, и в меньшей степени гор и пустынь. Даже в случае межконтинентальных заносов иксодид с мигрирующими птицами, регулярно происходящих между Европой, Африкой и Азией, мигранты обычно неспособны к закреплению в необычных для них геобиоценозах. Для ареалов иксодид характерно постепенное расширение или сужение границ в ответ на соответствующие многолетние изменения биотических и абиотических факторов, а не взрывная экспансия.
Исторические факторы в расселении иксодид отражают сложные процессы развития отдельных таксонов в условиях изоляции континентов и обмена фаун при соединениях континентальных платформ. В то же время на обширных массивах суши, как например Евразия или Северная и Южная Америка, распространение клещей определяется наряду с таковым их прокормителей абиотическими факторами среды и в первую очередь температурой и влажностью (табл. 1).
Наиболее морфологически примитивными и древними по происхождению среди Ixodidae считают в подсем. Ixodinae единственный род Ixodes, а в подсем. Аmblyomminae — роды Аропотта и Amblyomma. Эти 3 рода имеют космополитические ареалы, причем виды Ixodes распространились во всех климатических поясах, а два других рода — только в тропиках и субтропиках. По-видимому, предки этих родов существовали еще до распада единого массива суши — Пангеи на северную половину — Лавразию и южную — Гондвану. Это событие датируется серединой юры (Ушаков, Ясаманов, 1984; Оуэн, 1988), так что роды Ixodes, Аропотта и Amblyomma должны были существовать с середины мезозоя.
Дальнейшая судьба этих групп оказалась неодинаковой. Виды рода Аропотта (23) обитают преимущественно в разных типах тропических и субтропических лесов Восточного полушария (21 вид). Только в Новой Зеландии A. sphenodonti — паразит гаттерий — живет в умеренном климате, а в Австралии A. hydrosauri приспособился к аридным условиям пустынь. Наиболее богата фауна Азии (12 видов), в Австралии обитает 6 видов, в Африке — 4 и в Центральной и Южной Америке — 2 (рис. 26) (Колонии, 1983).
Род Amblyomma, включающий 102 вида, оказался более пластичным как в выборе прокормителей (рептилии, млекопитающие, птицы), так и по отношению к абиотическим факторам. Местообитаниями значительной части видов служат влажные тропические и субтропические леса, но большая часть африканских видов (A. variegatum, A. hebraeum, A. tholloni и др.) приспособилась к существованию в саваннах (Petney et al., 1987). В Южной и Центральной Америке также есть ксерофильные виды, встречающиеся в прериях (A. parvum, A. usingeri и др.). В зону с умеренным климатом заходят только A. americanum до 40° с. ш. в США (рис. 27) и A. parvitarsum, достигающий в Патагонии 50° ю. ш. Все остальные виды ограничены в своем распространении областями с тропическим и субтропическим климатом.
Наибольшее число видов (52) свойственно фауне Южной и Центральной Америки, включая Антильские острова и острова Галапагос с многочисленными эндемиками. Общие виды для Восточного и Западного полушарий отсутствуют, за исключением недавно завезенного из Африки на Антильские острова A. variegatum. В Африке обнаружено 19 видов, включая один эндемичный для Мадагаскара. В странах Южной и Юго-Восточной Азии встречается 17 видов и в Австралии — 12 (Колонии, 1983).
Самый многочисленный в семействе род Ixodes (около 250 видов) встречается во всех зонах вплоть до побережий полярных морей. Согласно Н. А. Филипповой (1977), внутри рода, включающего 19 подродов, имеются как более примитивные, так и эволюционно продвинутые группы. К числу первых принадлежат подроды Ceratixodes, Scaphixodes, Multidentatus, паразитирующие на древнейших по происхождению морских птицах, остатки которых известны с палеогена. Всесветные ареалы хозяев обусловили подобное же распространение их паразитов, не помогающее филогенетическим реконструкциям (рис. 28).
Из паразитов наземных млекопитающих наиболее морфологически примитивны подроды Sternalixodes, Exopalpiger, Endopalpiger, большая часть видов которых встречается только в Австралии, но по 1 виду подрода Exopalpiger встречается в Южной Америке, Африке и в Евразии. Судя по рецентным ареалам видов этого реликтового подрода, он должен был возникнуть в юре до распада Пангеи, и до настоящего времени эти клещи сохранили трофические связи с наиболее примитивными группами млекопитающих — однопроходными, сумчатыми и насекомоядными.
В пользу большой древности рода Ixodes свидетельствует и распространение наиболее богатого видами и эволюционно продвинутого подрода Ixodes s. str., отсутствующего только в Африке к югу от Сахары и в Австралии. По-видимому, этот подрод сформировался уже после распада Пангеи в ее северной половине — Лавразии до утраты наземного соединения Северной Европы, Америки и Европы, т. е. не позднее конца палеоцена. Дальнейшее расселение этих клещей шло в южном направлении на соединившийся с Азией в конце олигоцена — начале эоцена Индийский субконтинент, позднее в миоцене через Иберийский мост в Северную Африку, а из Северной Америки в Южную в плиоцене через образовавшийся тогда Панамский перешеек или же раньше, через острова Карибского моря. Последний путь менее вероятен, так как виды этого подрода не смогли преодолеть морские проливы между Южной Азией и Австралией.
Несмотря на широкое распространение, виды рода Ixodes повсеместно приурочены к местообитаниям с достаточной влагообеспеченностью. Виды, подстерегающие прокормителей на поверхности почвы или растительности, как например I. persulcatus и I. ricinus, приурочены к различным типам леса, где постоянно поддерживается высокая относительная влажность в приземном слое воздуха. В степях, саваннах и пустынях, где относительная влажность воздуха испытывает значительные сезонные и суточные колебания и может быть очень низкой, встречаются только виды с гнездово-норовым паразитизмом, который, по-видимому, возник независимо в нескольких подродах (Ixodes s. str., Afrixodes и др.). Примером подобных видов в Палеарктике служат I. laguri, I. subterraneus или I. осculms. Некоторые гнездово-норовые виды приспособились к жизни в околоводных норах с избыточным увлажнением, как например I. apronophorus в Палеарктике, I. ornithorynchi из нор утконосов в Австралии или I. aulacodi, паразитирующий в норах тростниковой крысы в Африке. Подобные же условия избыточной увлажненности характерны для местообитаний паразитов колониальных морских птиц — I. uriae, I. signatus и др.
Широкое распространение гнездово-норового паразитизма в роде Ixodes, вероятно, вторично, хотя и очень древнего происхождения. Ему должна была предшествовать стадия пастбищного паразитизма, обеспечившая выработку «жизненной формы» иксодового клеща как кровососа с чередованием стадий многодневного питания и еще более продолжительного свободного существования. На этой стадии эволюции должен был выработаться весь комплекс морфофизиологических и экологических адаптации, большинство из которых сохраняется при последующем переходе к гнездово-норовому паразитизму. Последний, несомненно, возникал неоднократно внутри разных эволюционных линий и встречается как в примитивных подродах (Coxixodes, Ceratixodes, Pholeoixodes и др.), так и в наиболее эволюционно продвинутом подроде Ixodes s. str. (I. apronophorus, I. laguri, I. occultus), в котором доминирует пастбищный тип паразитизма. В связи с этим следует отметить, что значительная часть видов морфологически примитивных подходов Exopalpige'r, Endopalpiger, Sternalixodes представлена не типичными гнездо-ю-норовыми, а пастбищными паразитами, как например австралийский I. (St.) holocyclus. У относящегося к этой группе I. (Ex.) trianguliceps, в частности, выявлен «примитивный» тип пастбищного паразитизма, при котором активные перемещения клещей ограничены самым верхним слоем почвы, растительной подстилкой и припочвенным ярусом растительности.
Второй по численности в семействе род Haemaphysalis (147 видов), согласно взглядам Хугстрала (Hoogstraal, Kim, 1985), включает 16 подродов, из которых 4 из 17 видов считаются эволюционно примитивными, а 12 со 130 видами — эволюционно продвинутыми. Подавляющая масса видов (более 90) обитает в Индо-Малайской области. В Эфиопской области встречается 19 видов и еще 12 эндемичных видов на Мадагаскаре, в Австралии — 12 видов, в Палеарктике — 19 и в Северной и Южной Америке — 4.
Большинство видов приурочено к разным типам зональных лесов тропической и субтропической зон. В умеренном климате при общем обеднении фауны эти клещи также встречаются в разных типах лесных местообитаний, причем могут иметь обширнейшие ареалы. Например, Н. concinna распространен от Западной Европы до Японии (рис. 33), а Н. leporispalustris от Аляски до Аргентины. Многие виды обитают в горных лесах. Общей особенностью всех этих местообитаний является достаточно высокая и стабильная влажность. Лишь немногие виды приспособились к существованию в аридных местообитаниях, как например H. рипctata, встречающийся в редколесьях и кустарниковых зарослях, или H. sulcata, встречающийся в степных и полупустынных горных районах Южной Европы, Передней и Средней Азии. Такие виды, как Н. erinacei или Н. hispanica из подрода Rhipistoma, обитают в ксерофильных ландшафтах, перейдя к гнездово-норовому паразитизму. На связь клещей рода Haemaphysalis с мезофильными лесными ландшафтами было впервые указано Б. И. Померанцевым (1948а), но вопрос о месте и времени происхождения и дальнейшей эволюции рода остается дискуссионным.
Хугстрал (Hoogstraal, Kim, 1985) предполагает, что род Haemaphysalis возник на Индостанском субконтиненте, откуда клещи в дальнейшем распространились в северо-западном направлении, достигнув побережий Атлантического и Тихого океанов. Они расселились по Африке, но лишь немногие смогли проникнуть в Северную и Южную Америку.
По нашему мнению (Балашов, 1993а), исходя из современных мобилистических представлений, подобная трактовка происхождения ареалов является спорной. Первоначально Индия была частью Гондваны, в мелу отделилась от нее и на протяжении многих миллионов лет оставалась изолированным островом. Столкновение Индии с Евразией произошло в конце олигоцена или начале эоцена и привело к мощным процессам горообразования, завершившимся образованием Гималаев. Гондванские корни рода представляются маловероятными, так как из других частей Гондваны Южная Америка, несмотря на обширную область тропических лесов, наиболее бедна представителями этого рода. Сравнительно небогаты фауны Haemaphysalis Австралии и Африки, причем на все 3 континента клещи несомненно проникли с севера. В связи со сказанным мы полагаем, что наиболее вероятны происхождение и ранние этапы эволюции рода в палеогене на территории Юго-Восточной Азии, где тропический климат и влажные леса сохранялись на протяжении всего кайнозоя. В пользу этой гипотезы свидетельствует распространение наиболее примитивных 3 из 4 современных видов подрода Allocerae в горных субтропических лесах этого региона, и только 1 — Н. inermis в лесах Южной Европы и Передней Азии. Первоначально эти клещи могли проникнуть в умеренно теплые части Евразии. Распространение клещей в Индостан и Африку произошло позднее, после их соединения с Евразией, а в Австралию они были занесены из Юго-Восточной Азии с птицами или, возможно, с грызунами сем. Muridae, сумевшими преодолеть морские проливы.
Из районов с тропическим и субтропическим климатом наиболее раннее вселение клещей произошло в Индию, и поэтому она богата видами этого рода по сравнению с другими осколками Гондваны. Крайняя бедность фауны Южной Америки может быть объяснена отсутствием клещей в фауне Северной Америки, куда они могли проникнуть через сравнительно холодную Берингийскую сушу и затем, двигаясь на юг через Панамский перешеек, попасть в Южную Америку. Эти события могли произойти только в плиоцене всего несколько миллионов лет назад, так что времени на образование эндемичной фауны оказалось недостаточно.
Очень обширным ареалом обладает род Dermacentor, насчитывающий около 32 видов (рис. 29). В Палеарктике обитает 15 видов, в Северной и Центральной Америке вплоть до 10° ю. ш. — 11 видов, в Тропической Азии — 4 вида и в Африке к югу от Сахары — 2 вида. Померанцев (19486) полагал, что этот род возник в горах или степях Центральной Азии и позднее по Берингийской суше проник в Северную и далее в Центральную Америку. Распространение этого вида в тропические области Старого Света происходило сперва с востока в Ориентальную область, а из нее в Эфиопскую. Согласно другой точке зрения (Белозеров, 1976; Колонии, 1984), палеарктические представители рода имеют североамериканское происхождение, а Евразийский центр видообразования этой группы вторичен.
Обе рассматриваемые гипотезы обходят стороной вопрос о хронологии описываемых процессов, но сходятся на единственно возможном пути расселения через Берингийскую сушу. Последняя возникала и исчезала неоднократно на протяжении кайнозоя, причем последние «мосты» датируются плиоценом-плейстоценом с возрастом в 3.5, 1.8 и 0.08 млн лет назад. Наиболее ранние мосты были возможны в конце мела или в плиоцене около 60 млн лет назад. Современные представления мобилистической геологии позволяют дать и третье возможное объяснение расселению рода Dermacentor, во многом сходное с таковым у хвостатых амфибий. Мы полагаем, что преимущественное распространение этих клещей в умеренных и умеренно теплых климатических зонах северного полушария и отсутствие в таковых южного допускают возникновение этих клещей в Лавразии уже после ее разделения с Гондваной, т. е. не ранее мела. В дальнейшем представители рода могли успеть расселиться по Евразии и Северной Америке еще до их расхождения и утраты соединения через Гренландию в палеоцене 45—50 млн лет назад, так что изоляция фаун этих континентов очень древняя. Широкому распространению клещей в южном направлении, скорее всего, препятствовали экологические факторы, и немногие виды, приспособившиеся к существованию в тропиках Юго-Восточной Азии и Африки, выделены систематиками в подроды Indocentor и Amblуocentor. Крайняя бедность фауны Южной Америки легко объяснима ее островным положением до плиоцена, так что немногие виды смогли проникнуть в этот регион не ранее 3 млн лет назад при массовом вселении в нее североамериканских млекопитающих через вновь образовавшийся Панамский перешеек. Возможно, что только D. nitens, иногда выделяемый в самостоятельный род Anocentor и широко распространенный в тропических и субтропических районах, мог проникнуть в Неотропическую область значительно раньше через острова Карибского моря вместе с ранними млекопитающими-вселенцами. Подобный путь распространения, в частности, допускается Траубом (Traub, 1980) для объяснения особенностей фаун блох и вшей этого региона.
Для палеарктических и большей части голарктических видов характерна приуроченность к различного рода степным или луговым ландшафтам, причем заметна связь многих из них с горным ландшафтом, а такие виды, как D. pavlovskyi или D. montanus, поднимаются в горах Средней Азии до верхних границ растительности на высотах 3500—4000 м над ур. м. (Филиппова, Панова, 1989). Встречающиеся в лесных ландшафтах клещи D. reticulatus и D. silvarum заселяют разного рода редколесья, вырубки, опушки и другие интразональные элементы. D. niveus заселил поймы рек с тугайной растительностью в аридной зоне. Можно предполагать, что связи этих клещей со степными и горно-степными ландшафтами, как предполагал Померанцев (19486), первичны, а с тропическими лесами — вторичны.
Из 73 видов рода Rhipicephalus не менее 55 встречается только в Африке, 8 — в Палеарктике и 4 — в Тропической Азии. В Австралии, Южной и Северной Америке, за исключением недавно завезенного R. sanguineus, клещи этого рода отсутствуют (рис. 30). Подавляющая масса видов заселяет разные типы степей, саванн и реже пустынь. В этих стациях клещи могут заселять интразональные редколесья и кустарниковые заросли, как например обитающий в тугаях R. leporis. Несколько видов обитает во влажных тропических лесах (R. cliffordi, R. complanatus, R. ziemanni). Гнездово-норовый паразитизм свойствен немногим видам (R. schulzei и R. pusillus).
Происхождение этого рода связывают с Африкой (Померанцев, 19486; Pegram, 1984), и эта гипотеза подтверждается геологическими данными. Известно, что с олигоцена Африка вместе с Аравийским полуостровом примерно 25 млн лет оставалась островом. Только 17 млн лет назад северо-восточный угол континента соединился через Аравию с турецкой и иранской частями Евразии, а 12—14 млн лет назад произошло соединение Африки с Иберийским полуостровом. Можно предполагать, что род Rhipicephalus сформировался в период островного существования Африки, а позднее, после соединения ее с Евразией, вместе с копытными и другими аборигенами африканской фауны распространился в аридные области Палеарктики. Там, в пустынях и степях Передней и Средней Азии, согласно Померанцеву (1948б), возник более молодой вторичный очаг видообразования.
Род Boophilus включает всего 5 видов, которые до завоза с домашним скотом в последние 200 лет отсутствовали в Австралии, Северной и Южной Америке. Предполагают (Hoogstraal, 1978), что первоначально распространение В. microplus было ограничено тропическими и субтропическими районами Азии, а в историческое время он проник в Америку, Африку и Австралию (рис. 31). В. kohlsi и В. annulatus происходят из Средиземноморья, а В. geigy и В. decoloratus — из Африки к югу от Сахары.
Главными прокормителями этих клещей повсеместно служат полорогие, и можно с достаточным основанием считать это семейство парнокопытных первичными хозяевами рода. Согласно палеонтологическим данным (Thenius, 1980), полорогие появились в Европе в раннем миоцене, но адаптивная радиация и расселение этого семейства, включая проникновение в Африку и Южную Азию, происходили в плиоцене. По-видимому, возраст рода, отличающегося высокой хозяинной специфичностью, не может быть старше, чем таковой его прокормителей.
Род Hyalomma включает около 30 видов, из которых примерно половина встречается в аридных местообитаниях Северной Африки, Южной Европы, Передней и Средней Азии (рис. 26). В пустыни Центральной Азии проникают только 3 вида этого рода. В саваннах и тропических редколесьях Африки к югу от Сахары обитает 5 видов и в сходных ландшафтах Индии — 5. Померанцев (19486) связывает происхождение рода Hyalomma с пустынями Передней Азии в эоцене, откуда эти клещи распространились в более мезофильные ландшафты Индии и Африки к югу от Сахары. Подобная точка зрения представляется более правдоподобной, чем взгляды Хугстрала (Hoogstraal, Kim, 1985) на возникновение этой группы еще в мелу. Независимо от мелового или более позднего происхождения их проникновение в Индию и Африку могло произойти не раньше неогена после соединения этих массивов суши с Евразией.