Иксодовые клещи — паразиты и переносчики инфекций
Видовой состав фауны иксодовых клещей в границах бывшего СССР
Видовой состав фауны иксодид в границах бывшего СССР достаточно разнообразен. Он насчитывает 86 видов (Филиппова, 1984), несмотря на нахождение территории в границах Палеарктики и зонах с умеренным или холодным климатом. Фауна России значительно беднее и включает около 60 видов, лишь немногие из которых имеют широкое распространение и достаточно многочисленны (см. приведенный ниже список). В Северной Евразии географические преграды распространению клещей отсутствуют. Однако существуют значительные различия в видовом составе фаун иксодид европейской части России и Дальнего Востока, разделенных пространствами Сибири с бедной фауной клещей. Видовые ареалы иксодид в наибольшей степени зависят от гигротермических условий окружающей среды и определяемых ими типов растительного покрова. Соответственно в широтном направлении с севера на юг разнообразие видового состава повсеместно увеличивается. Прослеживается четкая гвязь многих видов иксодид с определенными типами ландшафтов.
ВИДОВОЙ СОСТАВ ФАУНЫ ИКСОДОВЫХ КЛЕЩЕЙ (IXODIDAE) РОССИИ И СОПРЕДЕЛЬНЫХ ГОСУДАРСТВ
(по: Филиппова, 1984, с изменениями)
Amblyomma
1. A. gemma Donitz, 1909
Anomalohimalaya
2. A. lama Hoogstraal, Kaiser et Mitchell, 1970
3. A. lotozkyi Filippova et Panova, 1978
Boophilus
4.* B. annulatus Say, 1821 (B. calcaratus Birula, 1895)
Dermacentor
5. D. antrorum Resnik, 1950
6.* D. marginatus Sulzer, 1776
7. D. montanus Filippova et Panova, 1974
8.* D. niveus Neumann, 1897 (D. daghestanicus Olenev, 1929)
9.* D. nuttalli Olenev, 1927
10. D. pavlovskyi Olenev, 1927
11.* D. pomerantzevi Serdjucova, 1951
12. D. raskemensis Pomerantzev, 1946
13.* D. reticulatus Fabricius, 1794 (D. pictus Hermann, 1804)
14.* D.silvarum Olenev, 1927
15. D. ushakovae Filippova et Panova, 1989
Haemaphysalis
16. H. caucasica Olenev, 1928
17.* H. concinna Koch, 1844
18.* H. erinacei Pavesi, 1884 (H. numidiana Neumann, 1897)
19.* H. filippovae Bolotin, 1979
20.* H. flava Neumann, 1897
21.* H. inermis Birula, 1895
22.* H. japonica Warburton, 1908
23. H. kopetdagihca Kerbabaev, 1962
24. H. kutchensis Hoogstraal et Trapido, 1963
25.* H. longicornis Neumann, 1901 (H. neumanni Donitz, 1905)
26.* H. ornithophila Hoogstraal et Kohls, 1979
27.* H. parva Neumann, 1897 (H. otophila Schulze, 1918)
28. H. pavlovskyi Pospelova-Shtrom, 1935
29.* H. phasiana Saito, Hoogstraal et Wassef, 1954
30. H. pospelovashtromae Hoogstraal, 1966 (H. warburtoni Nuttall, 1912)
31.* H. punctata Canestrini et Fanzago, 1877
32.* H. sulcata Canestrini et Fanzago, 1877
33.* H. wellingtoni Nuttall et Warburton, 1908
Hyalomma
34.* H. aegyptium (Linnaeus, 1758)
35.* H. anatolicum Koch, 1844
36. H. asiaticum Schulze et Schlottke, 1929
37. H. detritum Schulze, 1919
38. H. dromedarii Koch, 1844
39. H. impressum Koch, 1844
40. H. kozlovi Olenev, 1934
41. H. kumari Sharif, 1928
42.* H. marginatum Koch, 1844
43. H. schulzei Olenev, 1931
44.* H. scupense Schulze, 1981
Rhipicephalus
45.* R. bursa Canestrini et Fanzago, 1877
46. R. leporis Pomerantzev, 1946
47.* R. pumilio Schulze, 1935
48.* R. rossicus Jakimov et Kohl-Jakimova, 1911
49.* R. sanguineus (Latreille, 1806)
50.* R. schulzei Olenev, 1929
51.* R. turanicus Pomerantzev, 194
Ixodes
52.* I. angustus Neumann, 1899
53.* II. apronophorus Schulze, 1924
54.* I. arboricola Schulze et Schlottke, 1929
55.* I. berlesei Birula, 1895
56. I. bruneus Koch, 1844
57.* I. caledonicus Nuttall, 1910
58. I. cornutus Lotozkyi, 1956
59.* I. crenulatus Koch, 1844
60. I. eldaricus Djaparidze, 1950
61.* I. frontalis (Panzer, 1798)
62. I. hexagonus Leach, 1815
63. I. kaiseri Arthur, 1957
64. I. kashmiricus Pomerantzev, 1948
65. I. kazakstani Olenev et Sorokoumov, 1934
66.* I. laguri Olenev, 1929
67.* I. lividus Koch, 1844
68.* I. maslovi Emeljanova et Kozlovskaya, 1967
69.* I. nipponensis Kitaoka et Saito, 1967
70.* I. occultus Pomerantzev, 1946
71.* I. pavlovskyi Pomerantzev, 1946
72.* I. persulcatus Schulze, 1930
73.* I. pomerantzevi Serdjukova, 1911
74.* I. redikorzevi Olenev, 1927
75.* I. ricinus (Linnaeus, 1758)
76.* I. sachalinensis Filippova, 1957
77. I. semenovi Olenev, 1929
78.* I. signatus Birula, 1895
79.* I. simplex Neumann, 1906
80.* I. strorqi Filippova, 1957
81. I. subterraneus Filippova, 1961
82.* I. trianguliceps Birula, 1895
83.* I. turdus Nakatsuji, 1943
84. I. unicavatus Neumann, 1908
85.* I. uriae White, 1852
86.* I. vesertilionis Koch, 1884
Примечание. * — виды, встречающиеся на территории России.
Дальше всех на север проникает I. uriae, заселяющий побережья Баренцева и Белого морей. Восточнее Новой Земли этот вид не проникает из-за суровости климата и снова появляется на побережье и островах морей Тихого океана. Клещи обитают в колониях морских птиц, часто образующих крупные скопления («птичьи базары»). В этих местообитаниях клещи заселяют многочисленные трещины и щели в скалах или же находятся в субстрате оснований гнезд. На островах Белого моря клещи встречаются в вырытых в земле гнездах чистиков. Местами численность клещей бывает очень высокой. На побережье Тихого океана встречается также клещ I. signatus, обладающий сходной экологией и часто встречающийся вместе с I. uriae. В зоне тундры иксодовые клещи отсутствуют.
В лесной зоне России достаточно часто встречается около 18 видов иксодид. Из них пастбищный тип паразитизма свойствен I. persulcatus, I. ricinus, I. pavlovskyi, I. nipponensis, D. reticulatus, D. silvarum, H. concinna, H. inermis, H. japonica, H. longicornis (Арзамасов, 1961; Филиппова,1977; Бойко и др., 1982; Успенская, 1987). Эти виды подстерегают своих хозяев, поднимаясь на растительность или же оставаясь на поверхности почвы. Наиболее широким распространением обладает таежный клещ I. persulcatus. Он встречается в среднетаежных, южнотаежных и широколиственных лесах от западных границ России до побережья и островов Тихого океана, включая юг Камчатки, а также в Северном Китае, Корее и Японии (рис. 32). В этих границах.клещи отсутствуют на заболоченных территориях, в сухих сосняках на песчаных почвах, а также на молодых вырубках и открытых лугах. На Сахалине в условиях высокой влажности воздуха клещи встречаются на закустаренных лугах и вырубках с высоким травостоем. Распространение клещей на север совпадает с изолиниями сумм температур воздуха 1400 °С за период с устойчивой среднесуточной температурой выше 10 °С или суммой температур 1600 °С за период со среднесуточной температурой выше 5 °С. Оптимальные же условия существования этого вида складываются при сумме температур более 2300 °С и коэффициентом увлажнения 0.45—0.60. В северной части ареала таежный клещ встречается только в ограниченных, наиболее теплообеспеченных местообитаниях. В европейской части ареала северная граница распространения вида проходит вблизи 62—63° с. ш., на Урале и в Сибири — около 60° и в Хабаровском крае между 52—54° с. ш. Южная граница ареала в европейской части России проходит от 56° с. ш. на западе до 53° на восток от Волги. В Сибири кроме зоны южной тайги клещи встречаются в изолированных лесных массивах лесостепи и по долинам рек и многочисленны в горных лесах Южной Сибири, Казахстана и Киргизии. На Дальнем Востоке южная граница ареала уходит на территорию Китая (Коренберг, 1979; Таежный клещ..., 1985). В горных лесах Южной Сибири и Дальнего Востока (рис. 32) вместе с таежным клещом встречается и другой лесной вид — I. pavlovskyi, отличающийся от первого вида преимущественным паразитированием взрослых особей на птицах.
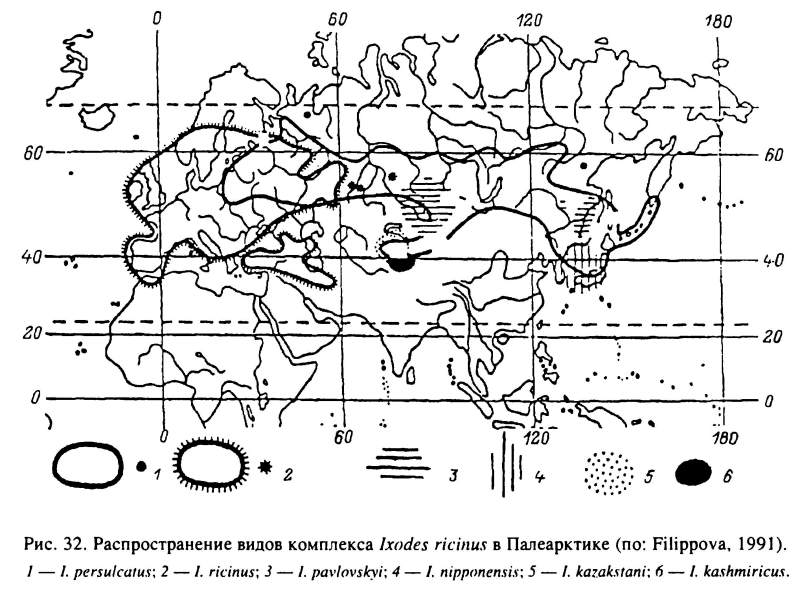
Ixodes ricinus распространен от Англии и Ирландии на западе до Волги на востоке (рис. 32). Во многих областях России к западу от Волги I. ricinus встречается вместе с I. persulcatus. Распространение этого вида на север ограничивается дефицитом тепла и совпадает с северной границей среднетаежных лесов. На юге клещи проникают за границы широколиственных лесов и многочисленны в зоне лесостепи и степи в разных типах искусственных лесонасаждений. Клещи также многочисленны в горных лесах Кавказа, Турции, Южной Европы и Северной Африки, где заселяют более влагообеспеченные леса на северных склонах и в речных долинах. Наиболее высокой численности этот вид достигает в зоне широколиственных лесов, но он достаточно многочислен и в расположенных севернее смешанных и южнотаежных лесах Скандинавского полуострова и России. I. ricinus легко приспосабливается к антропогенной трансформации ландшафта и заселяет разного типа вторичные леса среди сельскохозяйственных угодий (Померанцев, 1935). В условиях влажного климата Англии и Ирландии этот вид встречается не только в лесах, но также на открытых пастбищах, вересковых пустошах и в зарослях кустарников (Varma, 1965).
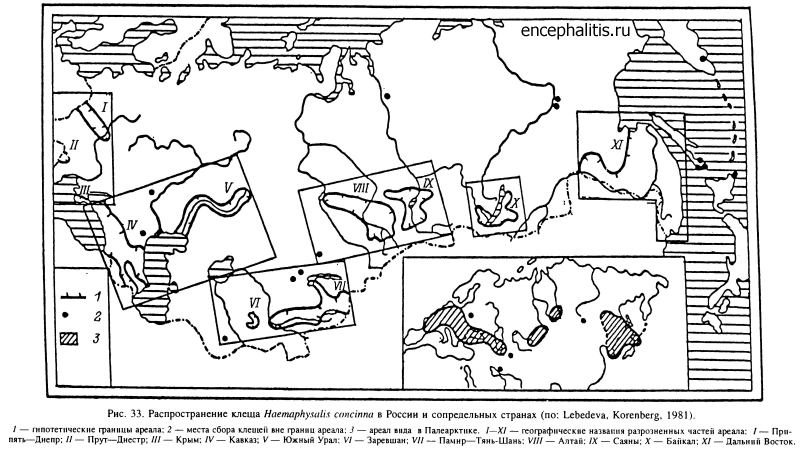
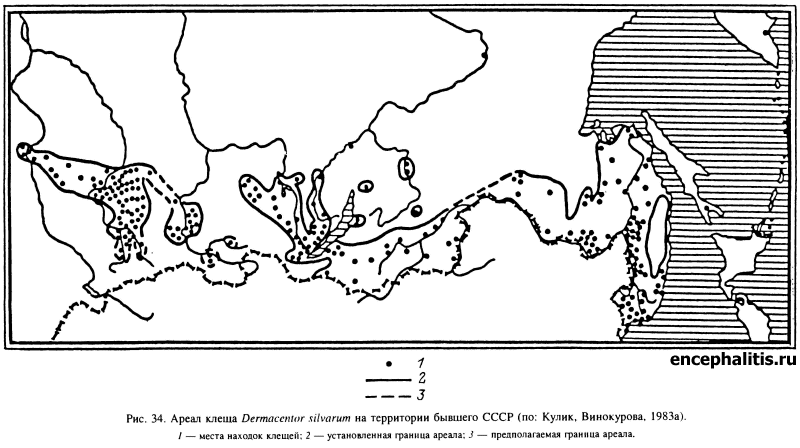
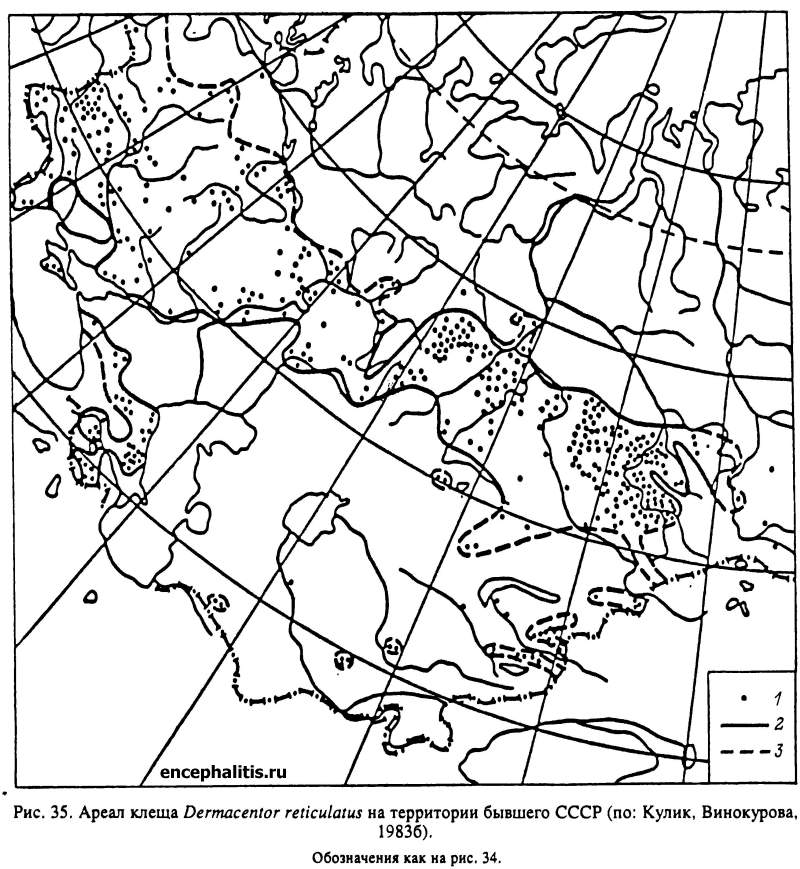
В широколиственных лесах Европы, включая юг России и Кавказ, обитают Н. inermis и Н. concinna. Второй из этих видов встречается также местами в лиственных лесах Тянь-Шаня, Алтая, Саян и многочислен в широколиственных лесах юга Дальнего Востока (рис. 33). В лесах последнего региона встречается еще несколько видов Haemaphysalis, из которых наиболее обычны Н. japonica и H. longicornis. На увлажненных луговых стациях прибрежной полосы в Южном Приморье встречается I. nipponensis, более многочисленный в Японии. В осветленных лесах, на лесных полянах, лугах и в кустарниковых зарослях по всему Дальнему Востоку и югу Сибири до Оби на западе распространен D. silvar (рис. 34). В зоне смешанных и широколиственных лесов Европы и Западной Сибири в луговых стациях, включая пойменные луга, в редколесьях и на вырубках многочислен D. reticulatus (рис. 35).
Гнездово-норовым типом паразитизма обладают клещи I. trianguliceps, I. apronophorus, I. crenulatus, I. lividus, I. arboricola, I. kaizeri, I. angustus, I. pomeranzevi. Клещ I. trianguliceps распространен в смешанных и широколиственных лесах от Англии до Енисея. Этот вид соединяет особенности пастбищного и гнездово-норового паразитизма. Клещи всю свою жизнь проводят внутри лесной подстилки в пронизывающих ее многочисленных ходах мелких млекопитающих. Голодные нимфы и самки этого вида могут подстерегать хозяев и на поверхности почвы, но не поднимаются на растительность. В лесах Дальнего Востока обитателями нор мелких млекопитающих являются I. angustus и I. pomeranzevi. В древесных гнездах птиц клещи встречаются только в зоне широколиственных лесов европейской части России и юга Дальнего Востока, где обитает относительно немногочисленный I. arboricola.
По поймам рек и заболоченным берегам озер от побережья Белого моря и до устьев рек юга России и Казахстана вплоть до Средней Сибири встречается I. apronophorus. Для этого вида характерно интразональное распространение, и его ареал охватывает лесную и степную зоны. Он заселяет норы связанных с водоемами главных прокормителей — водяной полевки и ондатры. Вторым интразональным видом является I. lividus, заселяющий гнезда ласточек-береговушек. Он распространен по всей Палеарктике от Атлантического до Тихого океана и по долинам рек встречается в полосе от 62° до 43° с. ш.
Видовой состав фауны клещей степной зоны значительно разнообразнее такового лесной зоны (Емчук, 1960; Миронов и др., 1965; Успенская, 1987). Часть из встречающихся в ней видов (I. apronophorus, I. lividus) отличается интразональным распространением, а в небольшие островки леса и искусственные лесонасаждения проникают с севера некоторые из типично лесных видов (I. ricinus, I. persulcatus, D. reticulatus, D. silvarum). Отличительной особенностью степной фауны является широкое распространение гнездово-норовых паразитов. От юга лесной зоны по всей степной зоне и горным степям Кавказа, Средней Азии и Южной Сибири в полосе от 54° до 35° с. ш. встречается I. crenulatus, заселяющий постоянные норы более 30 видов млекопитающих из числа хищников и сурков. В норах сусликов, хомяков и других грызунов на Украине, в южных областях России и в Западном Казахстане многочислен I. laguri. В норах сусликов и мелких хищников на юге европейской России, в Закавказье, Казахстане и Средней Азии часто встречается R. schulzei.
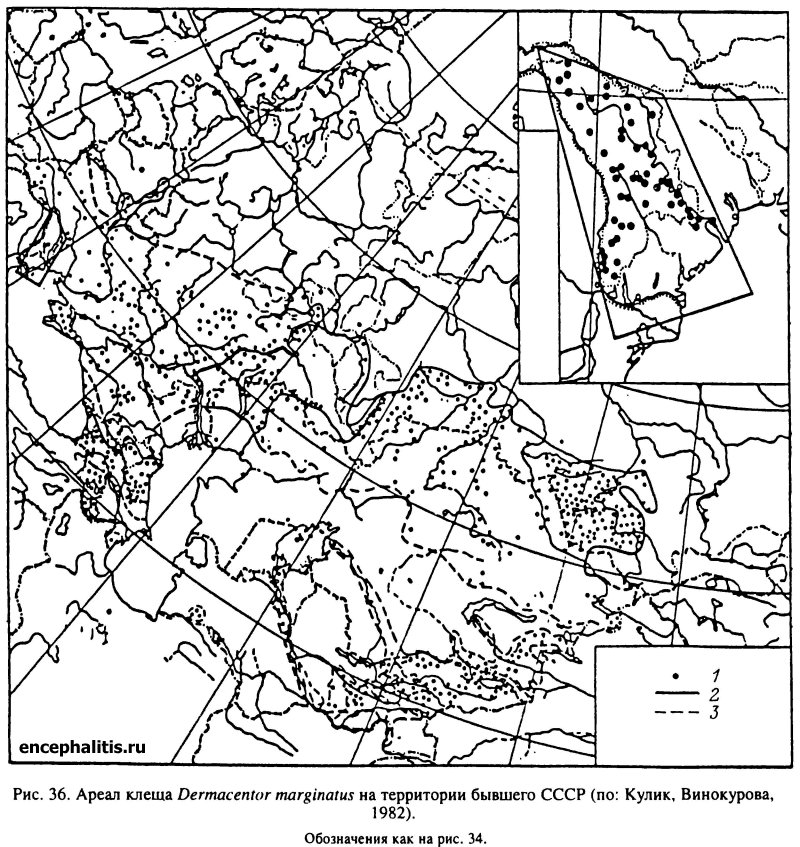
Характерными обитателями степей являются несколько видов с пастбищным типом паразитизма. В степных ландшафтах Европы, включая территорию России на восток до рек Томь и Катунь, обычен и многочислен D. marginatus (рис. 36). Этот же вид распространен в степных ландшафтах гор Кавказа и Средней Азии. В степях долины Енисея и особенно в Забайкалье обитает D. nuttalli, многочисленный также в Монголии и Северном Китае. В камышовых и кустарниковых зарослях пойм рек в аридных частях Ставропольского края, Дагестана, Закавказья, Средней Азии и Казахстана многочислен и часто встречается D. niveus. Из видов рода Haemaphysalis в степной зоне распространены H. punctata и Н. sulcata, особенно часто встречающиеся в горных ландшафтах. Из рода Rhipicephalus дальше всех проникает на север R. rossicus, встречающийся на Северном Кавказе, в Нижнем Поволжье, Северном Казахстане и Закавказье. В Саратовской, Ростовской и Астраханской областях, Ставропольском и Краснодарском краях, а также в Дагестане в степных и полупустынных ландшафтах встречаются Н. marginatum и Н. scupense. Особенно широко распространен первый вид, напитавшиеся нимфы которого заносятся далеко за границы ареала, от Поволжья и до островов на Балтийском море. Н. scupense паразитирует только на крупном рогатом скоте и имеет более ограниченное распространение.
Наиболее богата фауна южных степей, полупустынь и аридных предгорий Кавказского хребта (Ганиев, 1970). В Астраханской области и Дагестане появляется R. pumilio, более многочисленный в расположенных южнее пустынях и полупустынях Средней Азии и Казахстана. На овцах в Дагестане паразитирует двуххозяинный клещ — R. bursa, встречающийся также в Крыму, Краснодарском, Ставропольском краях. На Черноморском побережье Кавказа и Крыма на собаках паразитирует R. sanguineus. Только из Дагестана известны обычные для Закавказья и Средней Азии R. turanicus и Н. anatolicum, а в местообитаниях степной черепахи встречается паразитирующий на них H. aegyptium. В поймах и заболоченностях в Краснодарском крае и Дагестане местами встречается однохозяинный паразит крупного рогатого скота — В. annulatus. Для трех последних видов приводимые выше местонахождения в России являются крайними северными частями их ареалов. В целом для фауны степей характерно увеличение видового состава с севера на юг и обеднение с запада на восток.
В России полупустынные и пустынные ландшафты ограничены аридными районами Астраханской области, Калмыкии, Ставропольского края и Дагестана. Видовой состав фауны иксодид этих районов обеднен по сравнению с таковым южных степей. В расположенном южнее обширном поясе полупустынь и пустынь Казахстана и Средней Азии видовое разнообразие увеличивается в южном направлении и достигает максимума в пустынях Туркмении (Галузо, 1947, 1948, 1950; Бердыев, 1980). В закрепленных песках, заросших саксаулом, отмечено 13 видов. Из пастбищных паразитов доминирует Н. asiaticum, широко распространенный и в других типах пустынных ландшафтов. Он встречается от Восточного Закавказья и Ирана на западе до пустыни Гоби на востоке и проникает на север до Урало-Эмбинского междуречья в Казахстане. Реже в песках встречаются H. dromedarii и другие виды этого рода. Личинки и нимфы этого вида обитают в норах своих прокормителей — песчанок и других мелких млекопитающих, а взрослые — на верблюдах, крупном рогатом скоте, овцах и диких копытных. В норах грызунов и хищников встречаются I. occultus, I. redikorzevi, I. crenulatus, H. erinacei, R. schulzei. В глинистых пустынях Туркмении обнаружено 19 видов, многие из которых встречаются и в песках. Кроме Н. asiaticum на поверхности почвы здесь встречаются H. anatolicum и H. detritum, а на растительности — R. turanicus, Н. рипctata, H. sulcata, а из норовых видов многочислен I. subterraneus.
Богата фауна долин рек, протекающих по пустыне. Благодаря стыку пустынных ландшафтов с тугайными лесами, зарослями кустарников и камышей значительно возрастает разнообразие местообитаний, и в них встречается около 23 видов иксодид. Из них многочисленны Н. anatolicum, Н. dromedarii, Н. detritum, H. marginatum, В. annulatus, D. niveus, R. pumilio, R. turanicus.
Наиболее богата фауна клещей горных ландшафтов Закавказья и Средней Азии. Особенно разнообразна фауна гор Копетдага, насчитывающая 31 вид (Бердыев, 1980). Наряду с пустынными в ней много степных и горных видов. Особенно многочисленны в остепненных биотопах и стациях с кустарниково-древесной растительностью клещи R. turanicus, D. marginatus, D. niveus, H. sulcata, H. punctata, H. otophila, H. plumbeum, H. aegyptium, I. eldaricus, I. crenulatus. В местах концентрации крупного и мелкого рогатого скота обычны Н. anatolicum, H. detritum, R. bursa. В расположенных восточнее горных хребтах Памира и Тянь-Шаня вследствие большой высоты хребтов наблюдается большая вертикальная дифференциация фаунистических комплексов клещей (Гребенюк, 1966). В нижних поясах гор с пустынными и степными ландшафтами доминируют Н. marginatum, H. scupense, Н. anatolicum, H. asiaticum, В. annulatus, R. turanicus, H. sulcata. В зоне орехоплодовых лесов и кустарников многочисленны Н. punctata, D. marginatus, I. subterraneus, H. scupense, H. marginatum, R. turanicus. В еловых лесах, произрастающих на северных склонах и в ущельях Тянь-Шаня, доминирующим видом является I. persulcatus. В субальпийском и альпийском поясах на высоте 2500 м и более доминируют I. pavlovskyi, I. crenulatus, H. warburtoni.