Иксодовые клещи — паразиты и переносчики инфекций
Описания жизненных циклов клещей рода Dermacentor
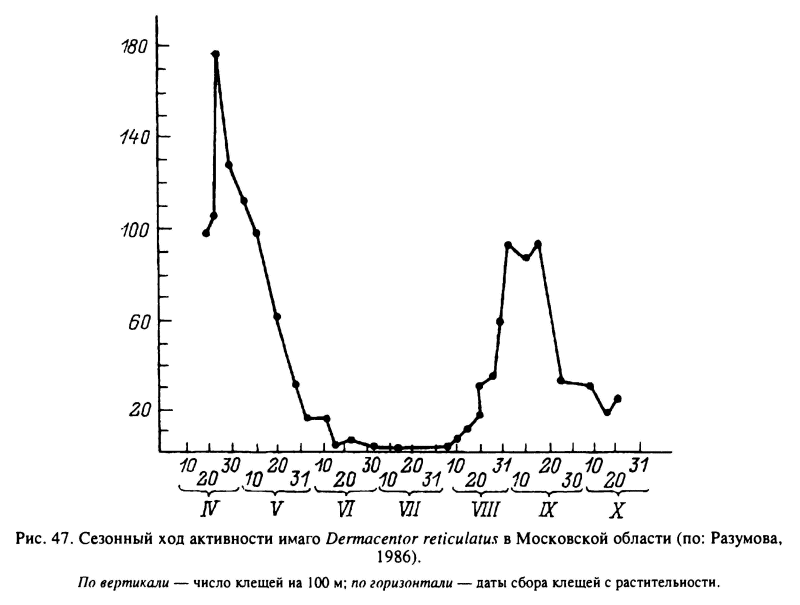
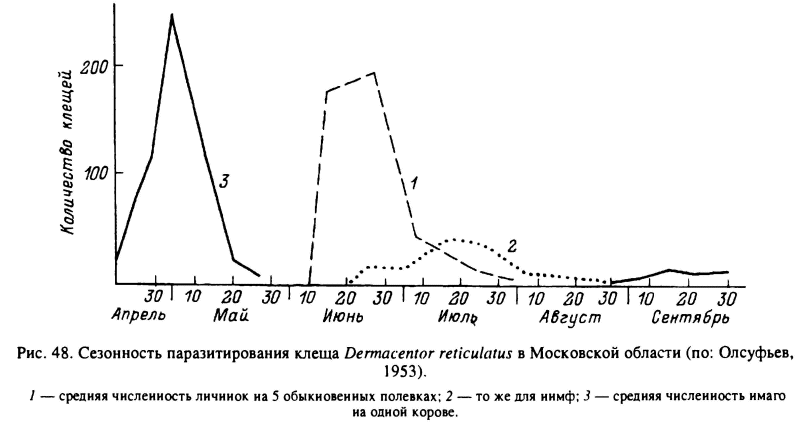
У клещей этого рода палеарктические виды характеризуются 1-годичными циклами развития, в сезонной регуляции которых главная роль принадлежит имагинальной диапаузе. Наиболее детально исследованы D. reticulatus (Олсуфьев, 1953; Daniel et al., 1980; Разумова, 1986) и D. marginatus (Покровская, 1951, 1953; Nosek, 1972; Gilot, Pautou, 1985), которые имеют во многом сходные циклы развития. D. reticulatus заселяет достаточно увлажненные луговые и кустарниковые биотопы в Европе и Западной Сибири, а ареал D. marginatus занимает зоны равнинных и горных степей. Повсеместно перезимовавшие голодные имаго этих видов активизируются со сходом снега ранней весной и нападают на скот, а также на средних и крупных диких млекопитающих до начала лета. В июне—июле клещи становятся неактивными и прекращают нападать на хозяев, хотя и могут оставаться на растительности. Нападение на скот возобновляется в августе—сентябре, но осенний пик активности значительно слабее весеннего (рис. 47, 48). У D. reticulatus в условиях Словакии и Польши весной в естественных местообитаниях этого вида самки приступали к откладке яиц в зависимости от температурных условий через 1—25 сут после насыщения, а личинки вылуплялись из яиц через 44—80 сут. Метаморфоз напитавшихся личинок занимал 12—22 сут и нимф — 25—60 сут. Голодные личинки живут не более 20—60 сут и нимфы — 35—70 сут (Daniel et al., 1980).
D. marginatus в условиях континентального и жаркого лета юга России обладает более сжатыми сроками развития. У самок весеннего поколения яйцекладка начинается через 9—12 сут, а вылупление личинок происходит через 12—15 сут.
Метаморфоз личинок занимает 3—7 сут и нимф — 22—26 сут. На все развитие от окончания питания самок до появления имаго следующего поколения требуется 90—110 сут. Голодные личинки живут в природе не более 30—40 сут и нимфы — 40—50 сут (Покровская, 1951, 1953). Из-за эфемерности неполовозрелых форм зимовка обоих видов возможна только на стадиях голодных имаго и напитавшихся самок, а голодные или напитавшиеся, но не перелинявшие к зиме нимфы погибают. Напротив, голодные имаго отличаются исключительной долговечностью. У D.reticulatus они могут жить в природе до 2.5 лет после линьки с 3 зимовками и 4 периодами активности, прерывающимися летом состоянием неактивности (рис. 49) (Разумова, 1966; Cerny et al., 1982).

Самки, напитавшиеся с середины лета, и осенью находятся в состоянии репродуктивной диапаузы и приступают к яйцекладке только после зимовки вместе с особями весеннего насыщения. Поведенческая и репродуктивная диапаузы регулируются длиной светового дня. Для D. marginatus характерна короткодневная фотопериодическая реакция с критическим фотопериодом в 13 ч (рис. 50) (Белозеров, 1976). Экологическое значение обеих форм диапаузы однозначно и направлено на предотвращение откладки яиц в конце лета и осенью и гибели не достигнувших имагинальной фазы особей зимой. Во время зимовки низкие температуры обеспечивают к весне реактивацию всего запаса зимующих имаго.

У клещей D. silvarum (Беляева, Рябова, 1971), D. nuttalli (Teng Kuo-fan, 1962; Меринов, 1964) и D. sinicus (Teng Kuo-fan, 1981), обитающих в Восточной Палеарктике, включая Северный Китай и Монголию, в связи с жарким летом развитие протекает еще быстрее, чем у видов из Западной Палеарктики. Перезимовавшие голодные имаго питаются на скоте, крупных и средних диких млекопитающих в апреле—июне. Их потомство благодаря быстрому метаморфозу напитавшихся летом личинок и нимф уже к осени дает самцов и самок следующего поколения, так что развитие одной генерации без зимовки занимает не более 3—4 мес. Например, у D. nuttalli (Teng Kuo-fan, 1962) напитавшиеся самки в условиях, близких к естественным, приступают к яйцекладке через 3—25 сут по отпадении, и она продолжается 31—44 сут. Развитие яиц занимает 15—36 сут. Личинки способны к питанию уже через 3—4 сут после вылупления и линяют через 8—12 сут после питания, а нимфы линяют через 14—20 сут. Осенний период активности у имаго выражен слабо. Иногда немногие прикрепившиеся к хозяевам особи остаются ненапитавшимися на протяжении зимы и заканчивают питание следующей весной. Подобную задержку прикрепившихся к хозяину самок D. silvarum в связи с их неспособностью к насыщению В. Н. Белозеров (1973) считает особой формой диапаузы, регулируемой на основе короткодневной фотопериодической реакции. Для самок D. sinicus также было показано удлинение сроков питания после нахождения голодных особей в течение 2.5 мес в условиях длинного дня и укорочение их после "Пребывания в условиях короткого дня (Jiang Zai-jie et al., 1991).
У D. auratus, одного из немногих видов, обитающих в тропических лесах Юго-Восточной Азии и Индии, жизненный цикл по ряду черт сходен с таковым палеарктических видов (Jiang Zai-jie, 1987). Имаго этого вида паразитируют на циких свиньях в летне-осенние и зимние месяцы. Особи из Южного Китая, напитавшиеся в июне—августе, приступают к яйцекладке через 13—27 сут, а в ноябре — через 215—244 сут, так что этому тропическому виду также свойственна репродуктивная диапауза (Jiang Zai-jie et al., 1991). Развитие яиц у этого вида более медленное по сравнению с северными видами (вылупление личинок через 45—79 сут), а сроки жизни голодных нимф больше (до 124 сут). В связи с диапаузой за 1 год успевает развиться одна генерация, но сроки активности всех фаз развития более растянуты и налегают друг на друга.
Жизненные циклы американских видов значительно разнообразнее евроазиатских как по продолжительности, так и по числу хозяев, сменяемых в развитии. Наиболее детально изучен D. variabilis, широко распространенный в США. Клещи заселяют мезофильные листопадные леса, кустарниковые редколесья и залежные луга. Имаго паразитируют на средних и крупных диких млекопитающих, собаках и скоте, а личинки и нимфы — на мышевидных грызунах, кроликах и зайцах. На юге (Newhouse, 1983) цикл развития 1-годичный. Зимуют голодные имаго (весенняя когорта) и личинки. Клещи активизируются уже в марте. Пик паразитирования личинок в апреле, а перелинявших из них и сразу же активизировавшихся нимф — в конце мая—начале июня. Перелинявшие из этих нимф имаго уже в конце июня—начале июля дают летний пик активности (летняя когорта). Напитавшиеся самки без задержки откладывают яйца, из которых вылупляются личинки, перезимовывающие в состоянии диапаузы. Потомство напитавшихся самок весенней когорты развивается с отставанием от особей летней когорты и уходит на зимовку на стадии голодных имаго.
На севере США в Массачусетсе (McEnroe, 1982, 1983) D. variabilis не хватает 1 года для завершения цикла развития. Напитавшиеся весной самки весенней когорты откладывают яйца, из которых в том же сезоне успевают вылупиться и напитаться личинки и перелинять в зимующих нимф. Эти нимфы питаются следующей весной и линяют в начале лета в имаго летней когорты. Последние успевают в этом же сезоне напитаться, откладывают яйца и оставляют зимующих личинок. Напитавшиеся весной личинки успевают перелинять в нимф, которые в свою очередь питаются и линяют в имаго весенней когорты, оставаясь неактивными до следующей весны. Значительная индивидуальная долговечность голодных нимф и имаго обеспечивает регулярную панмиксию между клещами весенней и летней когорт. У северной границы ареала в Манитобе (Канада) цикл развития уже только 2-годичный (Burachynsky, Galloway, 1985).
Экспериментально установлено (Smith, Cole, 1941), что личинкам и нимфам этого вида свойственна длиннодневная фотопериодическая реакция. Клещи реагируют не на саму длину дня, но на его сезонные удлинения или укорочения. Короткий день способствует сохранению неактивного состояния у уходящих на зимовку голодных личинок и нимф, а удлинение дня вызывает не только активацию, но и поддержание состояния активного подстерегания хозяина весной и в начале лета.
Клещ D. andersoni широко распространен в горных лесах и прилегающих к ним прериях системы Кордильер от Канады до Мексики. В зависимости от географической широты местности, высоты над уровнем моря и микроклимата местообитаний развитие возможно по 1-, 2- и 3-годичному циклам (Wilkinson, 1968, 1979). Имаго повсеместно имеют единственный весенний период активности с марта—апреля до начала июня. При наиболее распространенном 2-годичном цикле перезимовавшие имаго питаются весной, и из отложенных ими яиц в этом же сезоне вылупляются личинки. Последние, по-видимому, находятся в состоянии диапаузы и, не активизировавшись, зимуют. Следующей весной они нападают на мелких млекопитающих и после питания вскоре линяют в нимф. Нимфы также успевают напитаться и перелинять в имаго, которые неактивны осенью и зимуют. В случае зимовки не успевших напитаться голодных нимф развитие занимает 3 года, а при вылуплении личинок, а также питании личинок и нимф в один и тот же сезон развитие завершается за 1 год. Перезимовавшие, но не нашедшие весной хозяина имаго становятся неактивными, зимуют и вновь активизируются следующей весной.
Состояние активности у голодных особей D. andersoni стимулируется предварительной реактивацией низкими температурами и угнетается при воздействии температур более 25 °С. У «прерийной» расы этого вида (Pound, George, 1991) в условиях сокращающейся длины дня сроки развития напитавшихся нимф значительно удлиняются по сравнению с таковыми в условиях постоянного или удлиняющегося фотопериода. У напитавшихся самок из сокращающегося или постоянного короткого дня масса тела была больше, чем у особей из удлиняющегося или постоянного длинного дня. Сроки развития напитавшихся личинок достоверно не менялись при разных режимах освещенности. Для особой «горной» расы влияния длины дня на развитие не выявлено.
D. albipictus с однохозяинным развитием имеет 1-годичный цикл. Ареал этого вида охватывает Северную Америку от Канады до Мексики. Клещи паразитируют на лосях, белохвостом олене и в меньшей степени на других диких копытных, а также на лошадях и скоте. Личинки нападают на хозяев осенью. В Канаде (Альберта) в северной части видового ареала активные личинки появляются на растительности с начала сентября, их численность быстро нарастает до начала октября и затем постепенно сокращается до полного исчезновения в декабре (Drew, Samuel, 1985). Клещи встречаются на лосях осенью, зимой и весной, и минимальный период от прикрепления личинок до отпадения напитавшихся самок составляет 155 сут. Прикрепившиеся личинки сразу же начинают питаться и уже через 10— 22 сут появляются свежеперелинявшие нимфы. Последние остаются прикрепившимися, но не питаются в течение 3 мес до января, находясь в состоянии диапаузы. После этого они нормально питаются и линяют в имаго, которые также без задержки приступают к кровососанию. Количество имаго на лосях увеличивается с середины января до марта—апреля (Addison, McLaughlin, 1988).
Период отпадения с лосей напитавшихся самок приходится в Альберте на март—май. Яйцекладка начинается через 36—41 сут и занимает период с конца мая по конец июня. Личинки вылупляются через 76—106 сут (Drew, Samuel, 1986). После вылупления личинки остаются в скоплении в неактивном состоянии (поведенческая диапауза), которое в зависимости от географической широты местности продолжается от 4—7 мес в Калифорнии до 2—3 мес в Британской Колумбии и всего около 2 нед в Альберте (Drew, Samuel, 1985). Диапауза контролируется фотопериодической реакцией короткодневного типа, которая снимается осенью при сокращении светового дня до 12 ч (Wright, 1971).