Иксодовые клещи — паразиты и переносчики инфекций
Описания жизненных циклов клещей рода Ixodes
В наиболее богатом видами роде Ixodes при исключительно треххозяинном типе развития продолжительность существования каждой из фаз жизненного цикла и общая продолжительность последнего отличаются большим разнообразием.
В тропических и субтропических районах, по-видимому, достаточно широко распространены 1-годичные циклы. В частности, он свойствен обитателю лесов Восточной Австралии I. holocyclus. Активность личинок у него наблюдается летом-осенью, нимф — осенью-зимой и имаго — весной. У части популяции возможно удлинение развития до 2 лет (Doube, 1979). Сходный 1-годичный цикл существует и у I. rubicundus в Южной Африке (Fourie et al., 1989), причем у последнего вида активность ограничена достаточно влажными зимними месяцами. Как можно судить по последовательной смене периодов активности имаго, личинок и нимф в течение 1 года у I. matopi из Зимбабве, этот вид также должен иметь 1-годичный цикл развития, и он регулируется диапаузой у напитавшихся нимф (Colborne et al., 1981).
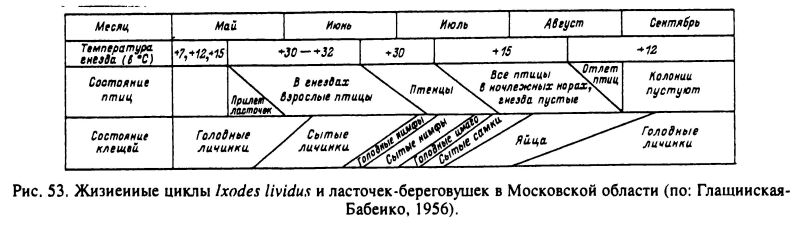
В умеренном климате 1-годичные циклы встречаются у обитателей нор и гнезд с достаточно теплым микроклиматом, но при этом часть популяций не успевает закончить развитие за теплый сезон и имеет 2-годичный цикл. I. lividus, паразит гнезд ласточек-береговушек (Riparia riparia), распространен по всей Палеарктике от Атлантического до Тихого океана. Быстрое развитие клеща в летние месяцы обеспечивается высокими температурами (30—32 °С) в гнезде хозяина в период насиживания яиц и развития птенцов. В Московской области (рис. 53) перезимовавшие в старых гнездах голодные личинки нападают во второй половине мая на прилетевших с юга взрослых птиц и питаются на них. Отпадение напитавшихся личинок происходит в конце мая—июне уже в новых гнездах. Нимфы появляются в конце июня—начале июля к моменту вылупления птенцов, сразу же нападают на них и отпадают внутри гнезда. Линька в имаго происходит в течение июля, и самки успевают напитаться на голых или уже оперившихся птенцах. Яйцекладка происходит с середины июля до начала августа. В конце августа—начале сентября вылупляются личинки, которые в состоянии диапаузы зимуют в оставленных птицами гнездах (Глащинская-Бабенко, 1956). В северной части ареала (Финляндия, Ленинградская область) небольшая часть популяции зимует на стадиях голодных нимф и имаго, и в этом случае цикл развития становится 2-годичным (Золотое, Букер, 1976; Ulmanen et al., 1977).
Сходным 1-годичным циклом, строго синхронизированным с гнездовым периодом хозяина, обладает клещ I. baergi — паразит скальной ласточки (Hirundo pyrrhonota) в южных аридных районах США (George, 1987; Larimore, 1987). В середине мая после прилета и начала гнездования ласточек голодные перезимовавшие личинки перемещаются из трещин в скалах внутрь гнезд, где нападают и питаются на взрослых птицах. Напитавшиеся клещи отпадают с хозяина, линяют в нимф, и последние присасываются к уже вылупившимся из яиц птенцам. Напитавшиеся нимфы выползают из гнезд, уходят в трещины скал и линяют в них во взрослых клещей. Голодные самки вновь перемещаются в гнезда и в первые две недели июля успевают напитаться на заканчивающих развитие птенцах. Отпавшие напитавшиеся самки выползают из гнезда, копулируют вблизи них с непитающимися самцами и уходят в трещины скал. В них происходит яйцекладка и вылупление личинок, которые остаются в неактивном состоянии до следующей весны. Как видно, в отличие от I. lividus у I. baergi развитие напитавшихся особей возможно вне гнезд ласточек благодаря высоким температурам юга США.
У клещей, обитающих в норах млекопитающих, благодаря возможностям контакта с хозяевами на протяжении всего года и более низким температурам внутри самой норы цикл развития менее жестко синхронизирован с таковым хозяина, и для его завершения может быть недостаточно 1 года. Так, у I. laguri — паразита сусликов рода Citellus, мелких хищников и других млекопитающих (свыше 30 видов), распространенного в степной зоне Евразии, все развитие протекает в норах хозяев. Зимуют голодные нимфы и имаго, которые активизируются весной и паразитируют на протяжении всего теплого сезона. Личинки от напитавшихся весной самок появляются в мае и тоже активны на протяжении лета—начала осени (Боженко, Шевченко, 1956). В условиях Чехословакии развитие одного поколения растянуто на 2 года (рис. 54), но часть популяции может развиваться и по 3-го-дичному циклу (Honzakova et al., 1980). В южных частях ареала возможен и 1-годичный цикл.

Сходным циклом развития обладает обитатель нор и гнезд околоводных млекопитающих I. apronophorus, распространенный по поймам рек и болот Европы и Южной Сибири. Главными прокормителями служат ондатра (Ondatra zibetica) и водяная полевка (Arvicola terrestris), а всего более 40 видов млекопитающих. В Молдавии (Успенская, 1987) в плавнях реки Прут самки, нимфы и личинки паразитируют с марта до декабря с максимумами в апреле-мае и августе-сентябре. У личинок осенний пик активности значительно выше весеннего. Зимуют голодные особи всех фаз развития. Цикл развития завершается за 1 год, но чаще он 2-го-дичный, а при отсутствии прокормителей может быть и 3-годичным.
Клещ I. crenulatus распространен в степной зоне Евразии и обитает в постоянных норах более 30 видов животных, из которых главными хозяевами служат несколько видов сурков (Marmota), барсуки, лисы, хорьки. Клещи паразитируют даже на некоторых видах птиц, контактирующих с норами млекопитающих. В условиях холодного климата в горах Тянь-Шаня (Афанасьева, 1951) в норах сурка развитие одного поколения занимает не менее 3 лет. Одновременно летом в гнезде встречаются все фазы развития клеща с 2 пиками численности на хозяевах. Первый пик дают перезимовавшие нимфы и самки в марте, а второй — в июне — свежеперелинявшие нимфы и личинки. В Молдавии (Успенская, 1987) в условиях более теплого климата клещи активны в норах барсуков, лис и других хищников на протяжении 10 мес. Пики численности питающихся самок приходятся на апрель, а нимфы многочисленны в течение всего теплого сезона с пиками в апреле и августе—октябре. Напитавшиеся весной личинки и нимфы успевают перелинять в этом же сезоне и напитаться в конце лета—начале осени. Личинки от летних кладок в основной массе остаются неактивными до следующей весны. По-видимому, для завершения развития 1 поколения в этих условиях требуется не менее 2 лет.
У видов с пастбищным типом паразитизма активность голодных особей и развитие напитавшихся протекает на растительности или на поверхности почвы. В результате эти клещи в большей степени зависят от местного климата и в первую очередь от гигротермических условий. В теплом умеренном климате для этих видов свойственны преимущественно 2-годичные циклы, но у части популяции наблюдается также 3-годичный цикл.
Преимущественно 2-годичным циклом обладает широко распространенный вдоль восточного побережья США I. scapularis (Yuval, Spielman, 1990). Личинки питаются на многих видах мелких млекопитающих и особенно на белоногом хомячке (Peromyscus leucopus) с июля по сентябрь. Перелинявшие из них нимфы активизируются после зимовки и также паразитируют на мелких млекопитающих с мая по июль. Перелинявшие в конце лета и осенью самки активизируются в этом же сезоне и большая часть популяции напитывается на белохвостом олене (Odocoileus virginianus) и других крупных и средних млекопитающих в осенние месяцы. При теплой погоде активность имаго продолжается зимой и заканчивается ранней весной следующего года. Существование осеннего и весеннего питания самок обусловливает разные сроки выплода и активации личинок и как следствие — наличие в популяции 2 частично обособленных когорт.
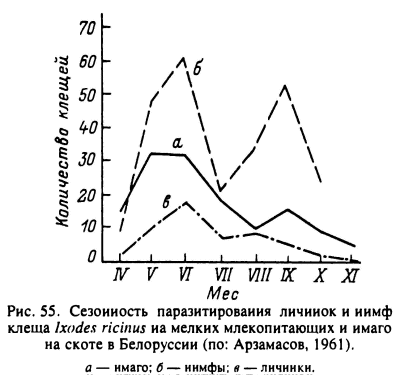
У некоторых видов, как например I. ricinus, жизненные циклы могут в зависимости от климата варьировать от 2- до 6-летних. В южных частях ареала, как например в горных лесах Алжира (Yousfi-Monod, Aeschlimann, 1986) или в Крыму (Мельникова, 1955), имаго активны в осенне-зимне-весенний сезоны непрерывно или с кратковременным прекращением во время сильных похолоданий. В умеренном климате Западной и Центральной Европы (Сегпу, 1957; Gigon, 1985; Gray, 1991; Craine et al., 1995), Белоруссии (Арзамасов, 1961) и средней полосы европейской части России (Бабенко, 1958) активность имаго приурочена к весенне-летне-осеннему сезону (рис. 55), причем весенний и осенний периоды обычно характеризуются максимумами численности, а летом может наступать снижение активности нападения клещей. В Карелии у северной границы ареала (Хейсин и др., 1955; Лутта, Шульман, 1958) при общем коротком периоде активности ее летний спад не выражен. У личинок и нимф в странах с умеренным климатом активность продолжается весь теплый сезон с разной степенью выраженности весеннего и осеннего пиков (Gray, 1991; Kurtenbach et al., 1995; Talleklint, Jaenson, 1996a).
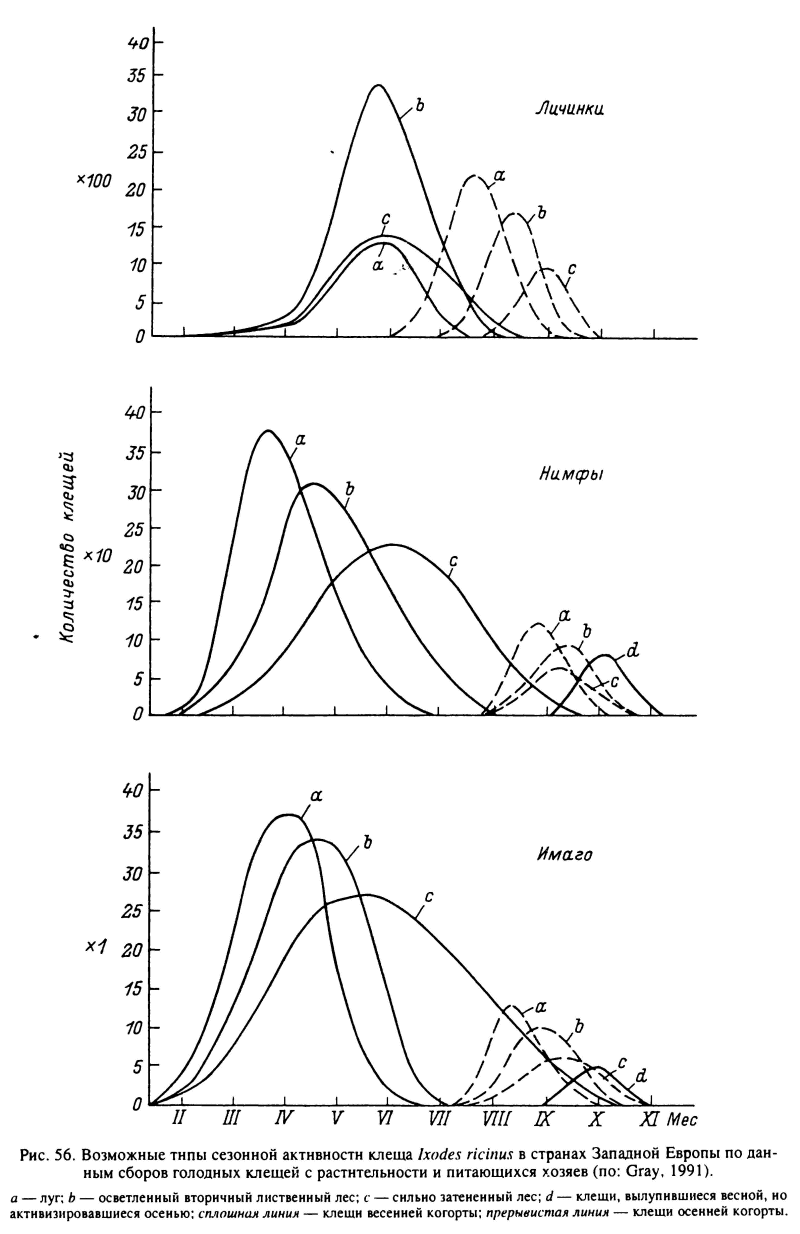
Сезонность активности у этого вида обеспечивается существованием морфогенетической диапаузы на фазах яйца, напитавшихся личинок, нимф и самок, так что выплод голодных особей происходит в основной массе во второй половине лета и осенью (Хейсин, 1955; Бабенко," 1958; Белозеров, 1988). Важную роль в синхронизации активности имеет также поведенческая диапауза у голодных особей. Она факультативна, контролируется фотопериодическими условиями и определяет соотношение между частями популяции, активизировавшимися в сезон вылупления или только после зимовки. Поэтому в южных частях ареала, как например в Молдавии (Успенская, 1987), осенний пик активности выражен значительно сильнее, чем на севере, в Карелии (Хейсин и др., 1955). Благодаря поведенческой диапаузе голодные особи на всех фазах могут переживать до 1 года и более, тогда как сроки жизни активизировавшихся клещей около 1 мес. Сроки активации перелинявших особей, продолжительность сезона их активности и индивидуальной жизни неактивных и активных особей определяются гигротермическими условиями в их местообитаниях и поэтому значительно варьируют в границах видового ареала (рис. 56).
Рассмотренные выше процессы в свою очередь определяют смену состава популяций, которые наряду с особями из разных когорт одной и той же генерации могут включать и особей, принадлежащих к 2 и более последовательным поколениям. Состав популяции I. ricinus, ее обилие и активность голодных особей обусловлены сезонностью развития яиц и напитавшихся личинок и нимф, сроками активации перелинявших особей и скоростью отмирания активных голодных клещей (Lees, Milne, 1951; Cerny, 1957; Бабенко, 1958; Балашов, 1962; Daniel et al., 1977; Gigon, 1985; Gray, 1991).
Для развития каждой из фаз жизненного цикла в большинстве случаев требуется не менее 1 года. Зимовка возможна на всех фазах развития в голодном и напитавшемся состоянии. ОДНАКО напитавшиеся клещи и яйца обязательно должны находиться в состоянии диапаузы, так как инициация осенью процессов эмбриогенеза или метаморфоза ведет к гибели клещей в зимний период. Благодаря факультативности диапаузы в южных частях ареала развитие при благоприятных условиях может завершиться за 2 года (Daniel et al., 1977). На севере, наоборот, для завершения 1 или всех фаз может потребоваться по 2 года, и у клещей наблюдался 5—6-летний цикл развития (Хейсин, 1955).
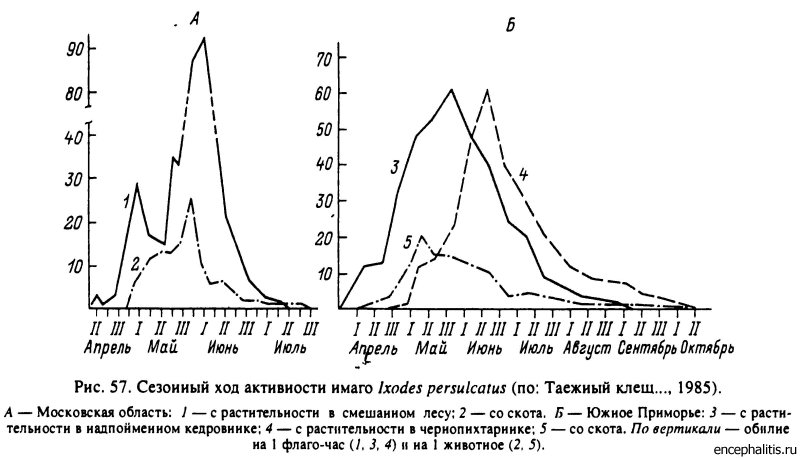
Сходным многолетним циклом развития обладает широко распространенный в таежных лесах Восточной Европы и Азии I. persulcatus. Активность голодных имаго наблюдается весной сразу со сходом снега и до середины лета (рис. 57). Они происходят от нимф, перелинявших осенью прошлого года, но остающихся в состоянии облигатной диапаузы в течение осенне-зимнего периода. Личинки и нимфы (рис. 58, 59) активны на протяжении всего теплого сезона и могут активизироваться сразу после вылупления из яиц или линьки во второй половине лета или же после зимовки как голодных, так и напитавшихся особей. У личинок и нимф диапауза факультативна, соответствует нормам длиннодневной фотопериодической реакции и предотвращает линьку особей, напитавшихся во второй половине лета. Напитавшиеся самки и отложенные осенью яйца неспособны к зимовке и погибают. Предполагают, что у голодных имаго поведенческая диапауза облигатна и предотвращает осеннюю активность клещей.


У японской популяции I. persulcatus (Fujimoto, 1993, 1995b) имаго, развивавшиеся в условиях длинного дня в первые 3 мес после линьки, находились в состоянии поведенческой диапаузы. В природе она снималась после зимовки к следующей весне. Длина дня в период от откладки яиц и до встречи с хозяином не вызывала поведенческой диапаузы у личинок. У голодных нимф, находившихся в условиях короткого дня, способность к питанию подавлялась. Режим короткого дня до, во время и после питания личинок тормозил линьку их в нимф, а в длинном дне развитие проходило без задержки. У напитавшихся нимф диапауза возникала при нахождении их в коротком дне до и во время питания. Условия содержания голодных нимф не влияли на их последующее развитие. Прочность поведенческой диапаузы голодных нимф и самок снижалась с увеличением времени, прошедшего после их линьки.
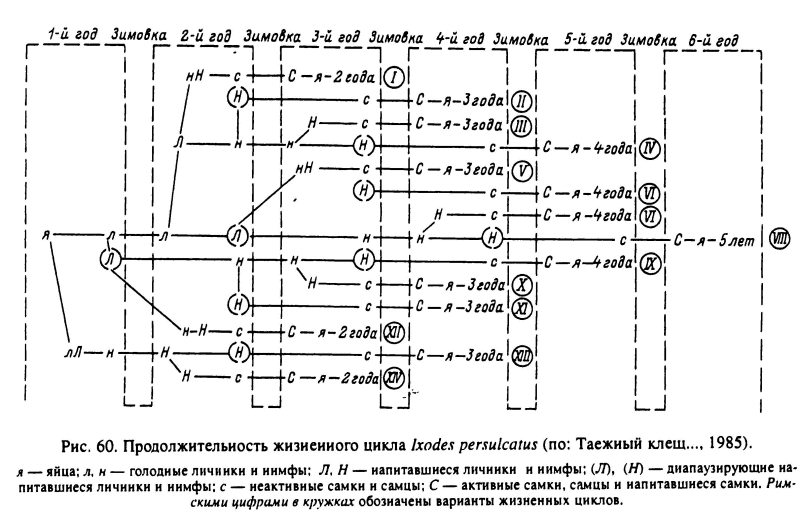
За 1 год обычно успевает закончить развитие только 1 фаза жизненного цикла, так что для завершения жизненного цикла требуется 3 года. В случае повторных зимовок на личиночной и нимфальной фазах, что нередко случается в условиях холодного лета и особенно у северных границ ареала, жизненный цикл 4- или 5-летний (рис. 60). В наиболее благоприятных условиях южных районов Дальнего Востока за теплый сезон успевают развиться яйца и вылупившиеся из них личинки и нимфы и развитие может уложиться в 2 года. Продолжительность жизненного цикла значительно варьирует не только в разных частях ареала, но и для особей одной и той же популяции (Таежный клещ..., 1985).
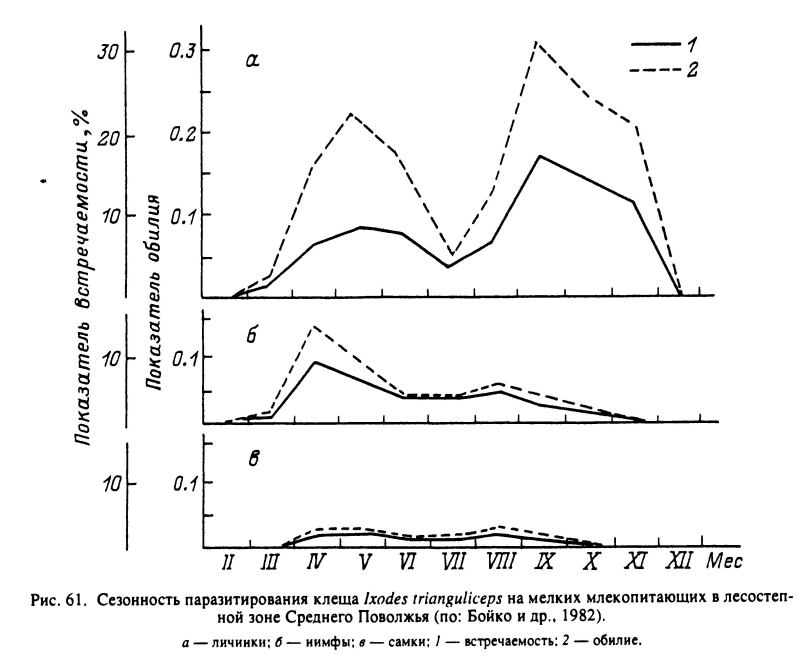
У I. trianguliceps, распространенного от Великобритании до реки Енисей на востоке в разных типах равнинных и горных лесов, продолжительность жизненного цикла может варьировать от 2 лет в Англии до 5 лет в Карелии. Активность всех фаз развития наблюдается круглогодично, включая и зимние месяцы (рис. 61). К зимовке способны напитавшиеся и голодные особи на всех фазах, а также яйца (Высоцкая, 1951; Бобровских, 1965; Randolph, 1975). В Швеции личинки паразитируют круглый год, кроме июля, с апрельским и сентябрьским пиками численности на прокормителях. Нимфы наиболее многочисленны на хозяевах весной, а летом и осенью их численность резко снижается. Самки также паразитируют круглый год с максимумом летом (Nilsson, 1974).
Дальше всех в приполярные области продвинулся паразит морских птиц I. uriae, и этому виду свойственны и наиболее продолжительные циклы развития. На побережье Баренцева моря развитие 1 поколения продолжается 5—7 лет (Карпович, 1973), в Ньюфаундленде — 4—5 лет (Eveleigh, Threlfall, 1974) и в более теплом климате у побережья Шотландии — 2—4 года (Barton et al., 1996). На птичьих базарах контакт клещей с птицами возможен только в гнездовой период длиною в 2—4 мес. В северных частях ареала каждая из активных фаз питается лишь 1 раз в году и их дальнейшее развитие из-за низких температур местообитаний может занять 2 летних сезона. Зимуют голодные и напитавшиеся особи. В Шотландии большая часть напитавшихся личинок и нимф линяет в том же сезоне, а немногие личинки майских сроков насыщения успевают напитаться, перелинять в нимф, и последние также питаются в этом же сезоне. За их счет часть популяции имеет 2-годичный цикл, но основная масса 3-годичный и меньшая часть — 4-го-дичный. Зимуют в подавляющей массе голодные особи всех фаз развития.