Иксодовые клещи — паразиты и переносчики инфекций
Синхронизация жизненных циклов с сезонными климатическими ритмами
Анализ данных о продолжительности и механизмах регуляции жизненных циклов у разных видов иксодовых клещей свидетельствует об их высокой лабильности. У видов с достаточно обширными ареалами в зависимости от климатических условий их местообитаний географические популяции могут различаться по таким важным параметрам, как фенология активности, зимующие стадии, механизм индукции диапаузы, время развития одного поколения. Продолжительность жизненного цикла у некоторых видов может варьировать даже для потомства одной самки, что создает исключительную возрастную гетерогенность клещевых популяций.
Оптимальные гигротермические условия для непрерывного развития иксодовых клещей в течение всего года, по-видимому, обеспечиваются влажным тропическим климатом. В субтропическом климате чрезмерно жаркий и сухой летний сезон или же холодный зимний могут замедлять или приостанавливать развитие, а в умеренном климате зимний перерыв в развитии иксодид становится правилом. Как и у насекомых, жизненные циклы иксодовых клещей приспособлены к особенностям климата их местообитаний. В условиях сезонных климатических ритмов периоды активной жизнедеятельности прерываются периодами покоя, особой формой которого является диапауза. На этой стадии у клещей повышается устойчивость к неблагоприятным факторам среды (низкие или высокие температуры, дефицит влажности) и благодаря существенному снижению уровня метаболизма удлиняются сроки жизни голодающих особей.
В. Н. Белозеров (1981) различает у иксодовых клещей 2 формы диапаузы. При морфогенетической диапаузе наблюдается задержка развития отложенных яиц, удлинение периода подготовки к линьке у напитавшихся личинок, нимф или оогенеза у самок. Поведенческая диапауза проявляется в отсутствии агрессивности (способности к нападению и питанию) у голодных клещей. Этот тип диапаузы может возникнуть сразу после вылупления или линьки и связан с уходом на зимовку, как в случае с имаго I. persulcatus и D. andersoni, или же с переживанием летнего сезона голодными личинками клещей Н. detritum или D. albipictus. В описанных случаях поведенческая диапауза накладывается на стадию послеличиночного до-развития. Последняя затягивается на несколько недель или месяцев, и для активации клещей необходимо сочетание определенных температурных условий и длины светового дня. У имаго D. reticulatus, D. marginatus и некоторых других видов, голодные имаго которых живут 2 года и более, поведенческая диапауза возможна после каждого периода активности и может повторяться в жизни одной особи 2 и даже 3 раза (Cerny et al., 1982; Разумова, 1986).
В согласовании во времени циклов развития иксодид с годовыми климатическими ритмами решающая роль принадлежит соотношению длины дня и ночи в суточном цикле (фотопериод). Реакцию организма на фотопериод, индуцирующую или предотвращающую развитие диапаузы, называют фотопериодической реакцией, и она обнаружена у многих видов иксодид (табл. 6). У клещей обнаружены длинные одневные фотопериодические реакции, когда активность или развитие стимулируется длинным днем, короткодневные реакции с противоположным эффектом и двухступенчатые реакции при реагировании на короткий и длинный день у нимф I. ricinus. Важной экологической характеристикой для организмов, обладающих фотопериодической реакцией, служит величина критического или порогового фотопериода, при которой соотношение особей с бездиапаузным развитием и диапаузой примерно одинаково.
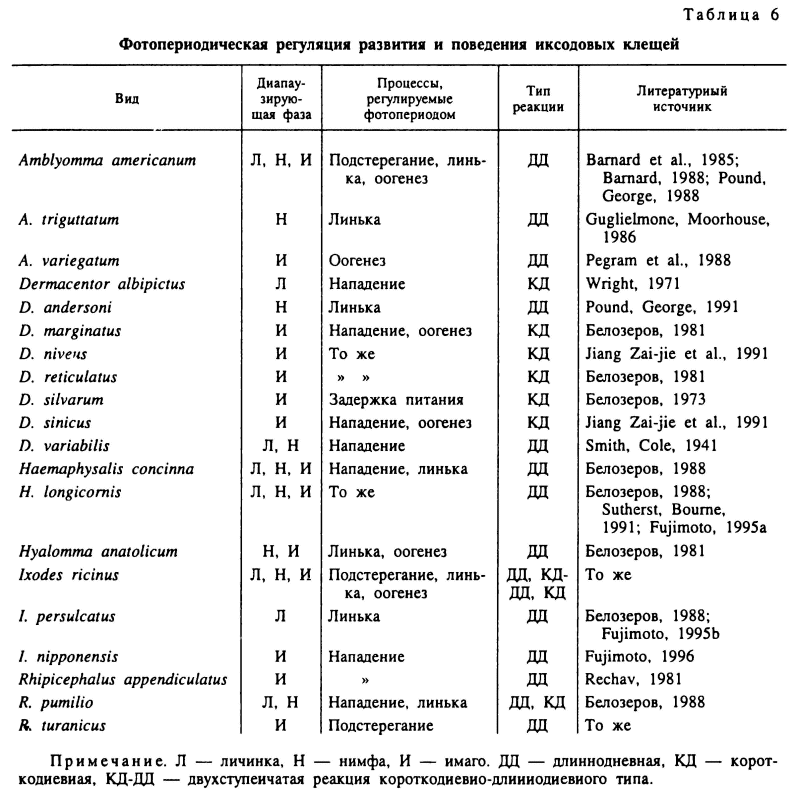
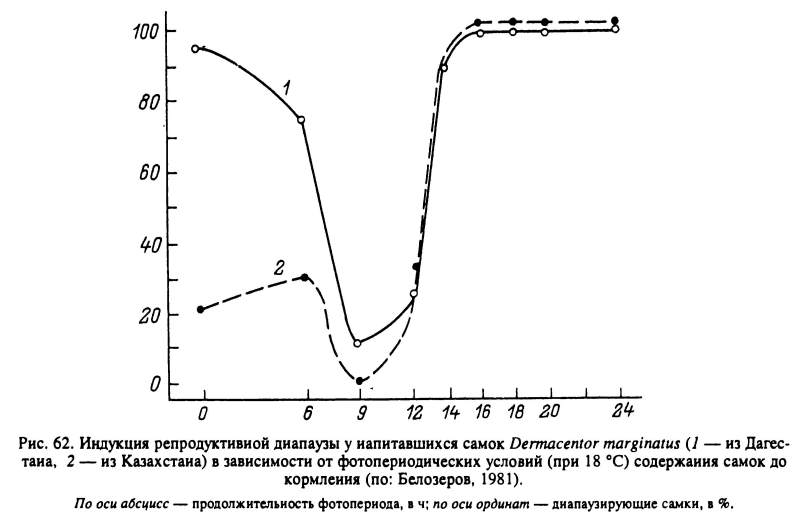
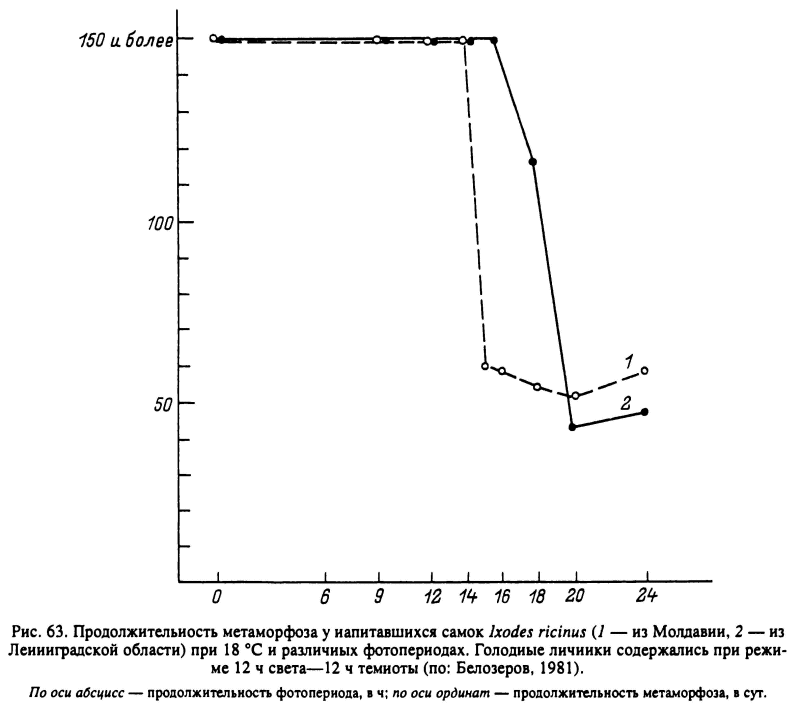
У клещей с длиннодневной фотопериодической реакцией, как например у нимф Н. anatolicum, при фотопериоде короче критического (13 ч) возникает диапауза. У видов с короткодневной фотопериодической реакцией, как например у напитавшихся самок D. marginatus, репродуктивная диапауза наступает при длине дня более 13 ч (рис. 62). Величина критического фотопериода может варьировать у разных географических популяций одного вида, увеличиваясь с юга на север. Например, у напитавшихся самок I. ricinus из Молдавии она составляет 15—16 ч света, а у личинок из Ленинградской области — 17—18 ч (рис. 63). Напротив, у D. marginatus этот показатель не был подвержен географической изменчивости и был равен 13 ч у популяций из Дагестана, Поволжья и Казахстана. При решающей роли фотопериода конечный результат фотопериодической реакции может зависеть от температуры (I. ricinus, Н. anatolicum), от возраста голодных особей (D. reticulatus, I. ricinus) и от других факторов (Белозеров, 1988).
Важной особенностью иксодид, обладающих многолетними циклами развития, является возможность повторного и независимого возникновения диапаузы на разных стадиях онтогенеза и в разные годы жизни клеща. Другой их особенностью является, в отличие от насекомых, частое совмещение на одной и той же стадии развития способности к восприятию фотопериода и ответа на него, как обнаружено у напитавшихся личинок и нимф I. ricinus или нимф и самок Н. anatolicum. Восприятие фотопериода на одной стадии развития, а реализация — на другой у иксодид встречаются реже, и примером этого могут служить голодные и напитавшиеся самки D. marginatus.
С возникновением диапаузы у клещей возникает повышенная устойчивость к неблагоприятным внешним условиям и особенно температуре. Наряду с понижением уровня метаболизма и экономным расходованием энергетических резервов организма клещ в этот период может проходить холодовую реактивацию. Последняя обеспечивает прекращение диапаузы в конце зимовки и стимулирует высокую активность перезимовавших голодных особей или яйцекладку и линьку у напитавшихся. Процесс холодовой реактивации достаточно длинен, и для ее завершения у напитавшихся самок D. marginatus при температуре 0—6 °С требуется 3—4 мес.
Наиболее полно исследована зимняя диапауза, характерная для иксодовых клещей, обитающих в регионах с достаточно холодной и продолжительной зимой. Стадии, на которых осуществляется зимовка, и факторы, индуцирующие диапаузу, в определенной степени видоспецифичны. Например, у I. persulcatus в пределах обширнейшего ареала этого вида самки, перелинявшие со второй половины лета, всегда остаются в состоянии облигатной диапаузы до весны следующего года. Напротив, у таких видов, как I. ricinus, постоянным признаком является лишь способность к возникновению диапаузы практически на всех стадиях развития. Конкретная же реализация этого признака определяется фотопериодом, метеорологическими условиями и физиологическим возрастом самих клещей. В результате реальное число стадий, на которых возникла диапауза, и общая продолжительность жизненного цикла могут быть различными не только для разных географических популяций, но даже для особей из потомства одной самки. Очевидно, что жизненные циклы иксодовых клещей представляют одну из важнейших адаптации к их условиям существования. По аналогии с насекомыми мы полагаем, что более простыми следует считать гомодинамные или, как их называют у клещей, «непрерывные» циклы развития, свойственные некоторым тропическим и субтропическим видам. Более сложные многолетние циклы с диапаузирующими стадиями мы рассматриваем как вторичные, возникшие в связи с приспособлением клещей к сезонным климатическим циклам умеренной зоны. Процессы, определяющие эволюцию жизненных циклов у клещей, исключительно лабильны и допускают возможность быстрого реагирования на флюктуации климата за счет перестройки жизненных циклов по их продолжительности, числу хозяев в развитии и срокам активности отдельных стадий.