Иксодовые клещи — паразиты и переносчики инфекций
Подготовка к яйцекладке и линьке
Напитавшиеся клещи, вследствие непропорционально больших размеров туловища по отношению к маленьким ногам, обладают ограниченной подвижностью. Они неспособны отползти от места отпадений от хозяина далее нескольких метров в поисках убежища. Их дальнейшая судьба поэтому в значительной степени зависит от отпадения в биотопах, благоприятных для дальнейшего развития. У многих видов это облегчается благодаря существованию суточных и иных ритмов отпадения особей, окончивших питание (табл. 7).
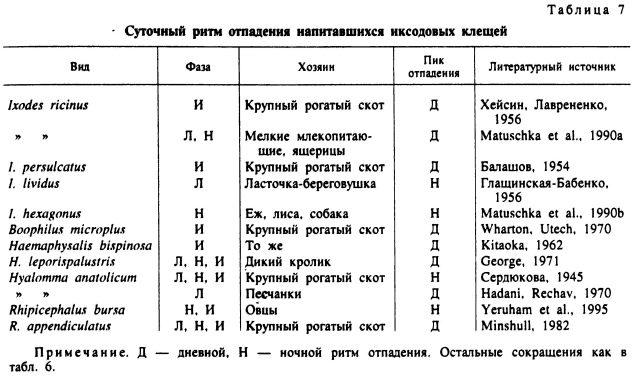
У гнездово-норовых видов иксодид ритм отпадения напитавшихся особей может быть приурочен ко времени нахождения хозяина в норе. Например, у H. leporispalustris (George, 1971) 90 % напитавшихся личинок, нимф и самок отпадали с диких кроликов в дневные часы, когда эти хозяева находятся внутри нор и других постоянных мест отдыха. Напротив, при паразитировании на ласточках-береговушках с дневной активностью напитавшиеся личинки I. lividus отпадали в ночные часы, когда хозяин находится внутри гнезда (Глащинская-Бабенко, 1956).
У пастбищных клещей время отпадения напитавшихся особей менее постоянно, вероятно в связи с большим разнообразием суточных ритмов активности их хозяев. У личинок и нимф I. ricinus, питающихся на грызунах и ящерицах, отмечены преимущественно дневной ритм отпадения напитавшихся особей и зависимость его от суточного ритма активности хозяев (Matuschka et al., 1990a). Напротив, у I. hexagonus, независимо от времени их прикрепления к главному прокормителю — ежу, напитавшиеся особи отпадают в ночные часы, когда активен их хозяин (Matuschka et al., 1990b). Самки клещей I. persulcatus (Балашов, 1954), I. ricinus (Хейсин, Лаврененко, 1956), В. microplus (Wharton, Utech, 1970) и все фазы R. арpendiculatus (Minshull, 1982) при питании на крупном рогатом скоте отпадают в утренние и дневные часы, когда скот выпасается на пастбищах и где в дальнейшем происходит яйцекладка и развитие личинок.
В условиях жаркого и сухого лета в Таджикистане личинки, нимфы и самки Н. anatolicum отпадали с крупного рогатого скота в ночные часы. В это время животные отдыхают в примитивных загонах, где и отпадают клещи. Дальнейшее развитие клещей происходит в трещинах глинобитных стен и под сухим навозом (Сердюкова, 1945). Подобный же, преимущественно ночной ритм отпадения свойствен нимфам и самкам R. bursa при питании на овцах в Израиле в летний период, а зимой значительная часть клещей отпадает и в дневные часы (Yeruham et al., 1995).
Напитавшиеся самки I. ricinus (Gigon, 1985) по оставлении хозяина в течение 1—10 сут сохраняют подвижность. За этот период они отыскивают укрытие под опадом листьев или внутри растительной подстилки. Радиус их поисковых перемещений ограничен 3—25 см и за день клещи могут проползти не более 5 см. После ухода под листья или в подстилку дальнейшее передвижение обычно прекращается, так как они неспособны к закапыванию в субстрат. Большая часть самок оставалась откладывать яйца в верхнем слое подстилки не глубже 3—5 см от ее поверхности.
Сходным образом вели себя и напитавшиеся I. persulcatus, хотя личинки и нимфы обладали несколько большей двигательной активностью. Нимфы и перелинявшие из них имаго расползались от места отпадения в радиусе до 5 м (Арумова, 1979). Расползающиеся личинки и нимфы иногда ненадолго поднимались на стебли травянистой растительности на высоту до 15—20 см. В конечном итоге они заползали в листовой опад или растительную подстилку на глубину в 1—3 см. Отпавшие самки также медленно перемещаются на расстояние в несколько сантиметров, пока не уйдут внутрь подстилки, мха или внутрь корней и трещин почвы. Отдельные особи были найдены в почве на глубине до 11 см, но активное зарывание в субстрат у этого вида также не обнаружено. У напитавшихся самок поведение определяется реакциями отрицательного фототаксиса, положительного тигмотаксиса и гигротаксиса (Таежный клещ..., 1985).
Для клещей, обитающих в аридном климате в открытых ландшафтах с разреженной растительностью, даже непродолжительное пребывание на освещенной солнцем поверхности почвы в дневные часы ведет к смерти от перегрева. У видов Hyalomma опасность перегрева уменьшается отпадением напитавшихся особей в ночные и ранние утренние часы. Отпавшие со скота напитавшиеся самки H. detritum сразу же стремятся уйти из-под прямых солнечных лучей под камни, в дерновины растительности, различные щели и полости в почве. При этом напитавшиеся особи способны проникать в очень узкие отверстия, меняя форму и диаметр своего туловища, но также неспособны к самостоятельному закапыванию в почву или песок (Галузо, 1947).
Время от момента отпадения напитавшейся особи до ее линьки на следующую фазу развития, а также подготовки к яйцекладке и самой яйцекладки значительно варьирует у разных видов, главным образом в зависимости от температуры. При бездиапаузном развитии (табл. 3, 4, 5) эти процессы занимают от нескольких суток до нескольких недель, а при диапаузе — до нескольких месяцев (рис. 50). Равным образом значительно варьируют и сроки развития яиц. С момента откладки яиц или после аполизиса клещи уже не могут перемещаться в поисках более благоприятных условий среды и целиком зависят от диапазона их колебаний. Успешность их развития в первую очередь определяется гигротермическим режимом в их микроубежище. При значительных видовых различиях в требованиях к условиям окружающей среды можно отметить значительную устойчивость к пребыванию в низкой относительной влажности (< 50 %) у видов Hyalomma, Rhipicephalus и Dermacentor и малую устойчивость у видов Ixodes и Haemaphysatis, многие из которых не могут развиваться при влажности менее 90 %. Наиболее чувствительны к дефициту насыщения отложенные яйца и наименее — напитавшиеся и готовящиеся к линьке нимфы, а также напитавшиеся самки.
Закончившие яйцекладку самки остаются живыми в течение нескольких суток, но в их организме продолжаются необратимые процессы распада кишечника и других внутренних органов, обязательно завершающиеся гибелью.