Иксодовые клещи — паразиты и переносчики инфекций
Холодостойкость
Для видов, обитающих в условиях умеренного и холодного климатов с достаточно суровой зимой, первостепенное значение может иметь холодостойкость зимующих стадий. К сожалению, данные о холодостойкости ограничиваются в основном лабораторными опытами с немногими видами. В этих экспериментах критериями холодостойкости служили температура или минимальная температура, при которой в течение определенного времени выживает определенная часть подопытных особей (табл. 12). Ввиду использования столь различных методик и разных стадий развития клещей приводимые далее фактические данные не вполне репрезентативны и позволяют установить лишь большую или меньшую степень холодостойкости в ряду изученных видов.
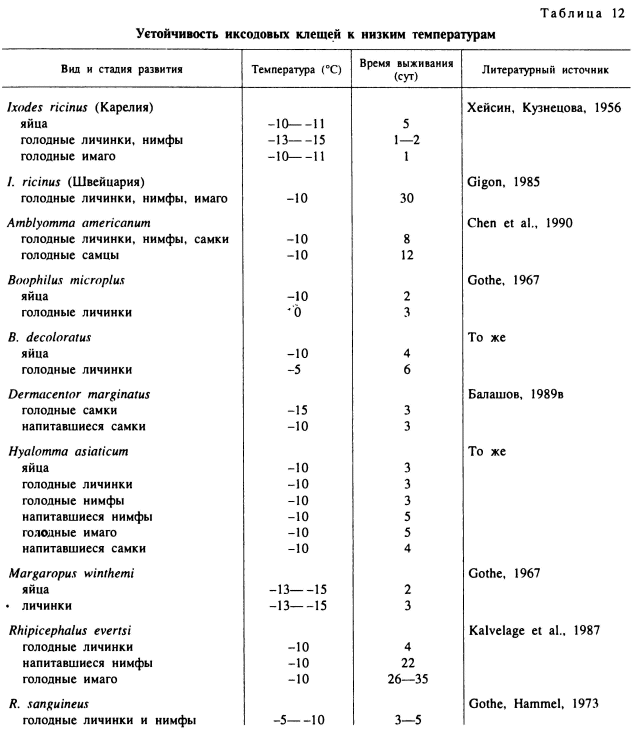
У обитателей умеренной климатической зоны — клещей I. ricinus и I. persulcatus у их северных популяций из Карелии в лабораторных опытах все стадии развития выживали 7 и более суток при температурах до -10 °С, а более низкие температуры (до -15 °С) вели к гибели клещей через 1—3 сут (Хейсин, Кузнецова, 1956). Близкие результаты получены и для популяции I. ricinus в более теплом климате Швейцарии. Голодные и напитавшиеся личинки, нимфы и имаго этих клещей без вреда переносили по несколько повторных суточных экспозиций при -4.5 °С (рис. 71), тогда как при -10.5 °С смертность значительно увеличивалась (Gigon, 1985). Достаточно высокая холодостойкость была обнаружена у личинок и нимф I. persulcatus из Японии, причем у голодных особей она была несколько выше, чем у напитавшихся. Нахождение от 1 до 50 сут при температуре -2 °С вообще существенно не влияло на смертность голодных особей и способность к линьке у напитавшихся. Через 10 сут пребывания при температуре -5 °С выживали 100 % голодных личинок и 50 % нимф, а через 50 сут — 37.5 % личинок и 80 % нимф. Среди напитавшихся личинок через 30 сут пребывания при -5 °С в дальнейшем перилиняли 40 % личинок и 20 % нимф.
Достоверные различия в холодостойкости диапаузирующих и недиапаузирующих напитавшихся клещей не выявлены. Холодостойкость яиц была значительно ниже, и после их нахождения при 4 °С или -2 °С в течение от 30 сут вылупление личинок произошло соответственно в 29 и 4 % случаев. Через 50 сут пребывания при подобных низких температурах погибли 99 и 100 % отложенных яиц соответственно (Fujimoto, 1994, 1995с).
В условиях мягкого климата Западной и Центральной Европы температура в местообитаниях зимующих I. ricinus (растительный опад и верхний слой почвы) большую часть зимы остается выше 0 °С и лишь периодически опускается до -2 °С (Daniel et al., 1972; Gigon, 1985). Очевидно, что присущая этим клещам холодостойкость обеспечивает их успешную перезимовку.
В Восточной Европе и Сибири у северных границ ареалов I. ricinus и I. persulcatus зимние максимумы температур достигают -30 и даже -40 °С. В этих условиях перезимовка клещей возможна только под снежным покровом. Например, при температуре воздуха -30 °С под слоем снега в 25—30 см температура поверхности почвы не опускается ниже -6 °С, а под слоем снега в 75 см — не чиже -1 °С. Сопоставление показателей холодостойкости клещей и максимальных значений температур в местах их зимовки свидетельствует не только о высокой степени приспособленности этих видов к переживанию зимнего периода, но и о наличии у них определенного «резерва холодостойкости» на случай экстремальных температур при отсутствии снега в начале зимовки.
У клеща D. variabilis, северная граница распространения которого в Северной Америке в основном совпадает с январской изотермой -1 °С (Sonenshine, 1979), холодовая устойчивость оказалась несколько меньшей. Среди голодных личинок этого вида, находившихся в течение 24 ч при температуре 0 и -2 °С, выжили соответственно 47.8 и 21.9 %, а среди голодных взрослых — 38.5 и 3.9 % (Smith et al., 1946). У северной границы распространения этого вида установлена прямая зависимость выживания голодных самок от температурного режима и наличия постоянного снежного покрова (McEnroe, 1984, 1990). В Массачусетсе успешная перезимовка голодных клещей возможна при установлении устойчивого снежного покрова до наступления морозов, так что клещи находятся под защитой от высыхания и температур ниже 0 °С. В результате аномально холодной зимы, какая наблюдалась в 1979—1980 гг., когда морозы начались и держались до выпадения снега, значительная часть зимовавших голодных нимф погибла и численность их весной и летом была крайне низкой. Для восстановления прежней высокой численности потребовалось 2 года с благоприятными климатическими условиями.
У широко распространенного в степных местообитаниях Европы и западной части Азии D. marginatus единственной зимующей фазой являются голодные или напитавшиеся имаго, переносящие в течение 3 сут охлаждение до -15 °С (табл. 12), что предотвращает вымерзание клещей в зимние месяцы. Для успешности перезимовки этих клещей в особенно суровом климате горной Армении (Рухкян, 1986) важную роль играет толщина снежного покрова. В местообитаниях с большей толщиной снежного покрова процент перезимовавших самок был выше, чем в стациях, где снег сносится ветром.
Крайне суровы условия зимовки для клещей — обитателей северных пустынь Казахстана и Средней Азии. Зимние минимумы достигают здесь -20 °С и часто наблюдаются при отсутствии снежного покрова. В Южном Казахстане в микробиотопах местных видов Hyalomma минимальные температуры припочвенного слоя воздуха опускаются от -5 до -10 °С. Клещи избегают промерзания благодаря зимовке в примитивных глинобитных постройках для скота, как в случае с Н. anatolicum, где температура обычно выше 0 °С (Галузо, 1947). Наиболее же часто клещи зимуют внутри подземных нор большой песчанки, как в случае с Н. asiaticum, а также других грызунов и хищников. В зависимости от глубины норы зимние температуры в подобных микробиотопах около 0 °С и более. В наших опытах (табл. 12) все стадии развития Н. asiaticum без вреда для последующего развития переносили пребывание в течение 3 сут при температурах от -5 °С (личинки) до -15 °С (напитавшиеся нимфы). Подобная холодостойкость «с запасом» гарантирует успешную перезимовку этих клещей в норах обычной песчанки.
Субтропические виды иксодид также успешно переносят зимние понижения температуры ниже 0 °С. Например, голодные личинки и нимфы R. sanguineus в течение нескольких дней выдерживали охлаждение до -5 °С, а напитавшиеся нимфы — 1 сут при -10 °С, 3 сут при -5 °С. Голодные имаго переносят пребывание при -10 °С до 6 сут (Gothe, Hammel, 1973). Кратковременные понижения температуры до 0 °С и ниже не причиняли вреда зимующим в условиях Иерусалима голодным имаго и напитавшимся нимфам этого вида (Feldman-Muhsam, 1981). У A. americanum значительная часть голодных имаго, нимф и личинок выдерживала 8-суточное пребывание при температуре -10 °С. Холодостойкость личинок и нимф была ниже, чем имаго, а напитавшихся особей ниже, чем голодных. Экспозиция в течение 7 сут при -17 °С вызывала 100 %-ную гибель клещей (Chen et al., 1990; Needham et al., 1996).
Холодостойкость изученных субтропических видов клещей во всех случаях оказалась большей, чем минимальные температуры, наблюдающиеся в их местообитаниях. Примером служит R. evertsi mimeticus, который распространен в странах Южной и Восточной Африки, где зимние температуры выше 0 °С. Тем не менее этому виду свойственна довольно высокая холодостойкость. Голодные имаго выдерживали пребывание при -10 °С от 3 сут в условиях 15 %-ной относительной влажности, до 11 сут в 100 %-ной относительной влажности, а напитавшиеся нимфы при этой же температуре выживали даже 22 сут (Kalvelage et al., 1987).
В других случаях прослеживается связь холодостойкости с климатическими особенностями местообитаний клещей. В Южной Африке встречаются два вида — В. microplus и В. decoloratus, из которых первый завезен из тропических районов Юго-Восточной Азии, а второй — абориген. Наименьшая холодостойкость обнаружена у В. microplus, голодные личинки которого погибают уже после 3-суточного пребывания при 0 °С, тогда как у В. decolorants они выживают до 6 сут при -5 °С (табл. 12). У Margaropus winthemi — однохозяинного клеща, обитающего в горных районах Южной Африки, холодостойкость не уступает обитателям умеренной зоны. Яйца и личинки этого клеща выдерживали в течение 2—3 сут температуры от -13 до -15 °С (Gothe, 1967).
Сведения о биохимических и физиологических механизмах холодостойкости для иксодовых клещей ограничены. Сопоставление разрозненных данных о температурах переохлаждения и сроках выживания этих клещей при минусовых температурах позволяет сделать вывод, что иксодиды погибают, когда температура опускается ниже точки переохлаждения й в их организме образуется лед. Это свойственно даже I. uriae, который зимует в верхних слоях почвы, неглубоких трещинах и щелях в скалах на птичьих базарах в Арктике и Антарктике, где возможны кратковременные понижения температуры воздуха до -30 °С. Температура переохлаждения у напитавшихся личинок составляет от -18.9 до -20.9 °С, у нимф — от -11.5 до -19.5 °С и самок — от -7.1 до -12.7 °С. Самая низкая температура переохлаждения обнаружена у яиц и составляет -28.7 °С (Lee, Baust, 1987).
У распространившегося на север до Южного Казахстана клеща Н. detritwn температура переохлаждения напитавшихся нимф варьирует от -15 до -18 °С. У свеженапитавшихся нимф Н. anatolicum она не превышает -10 °С и нимф в конце метаморфоза -18 °С, а у голодных имаго — от -6.5 до -12 °С (Галузо, 1947).
Интересно, что у клеща A. americanun, обитающего в условиях теплого умеренного климата Северной Америки, температура переохлаждения для разных стадий развития существенно не отличается от таковой для клещей из регионов с более суровым климатом. У имаго A. americanum этот показатель составил -13 °С, у напитавшихся нимф — от -16 до -18 °С, у напитавшихся личинок -22 и яиц -27 °С (Chen et al., 1990; Needham et al., 1996).
Очевидно, что всем иксодидам свойствен единый механизм устойчивости к низким температурам. Клещи выживают только при температурах выше точки переохлаждения. Более низкие температуры вызывают образование льда в их организме и гибель. Несомненно, чтсг, несмотря на близкие нижние границы температур переохлаждения у разных видов, определяющиеся осмотическим давлением гемолимфы, существуют достоверные различия в способности к переживанию при низких температурах не только у разных видов, но и у разных стадий развития одного вида. Физиологические механизмы подобных различий не исследованы, и имеются лишь указания на изменения в осмотическом давлении гемолимфы и содержании в ней глицерина, понижающих точку переохлаждения (Lee, Baust, 1987; Chen et al., 1990).