Иксодовые клещи — паразиты и переносчики инфекций
Потери воды организмом
Ввиду первостепенного значения влажности окружающей среды в экологии иксодовых клещей различные аспекты этой проблемы рассмотрены для многих видов. Специальные исследования посвящены также механизмам регулирования водного баланса у иксодид, и эти вопросы рассмотрены в нескольких обзорах (Knulle, Rudolph, 1982; Балашов, 1989в; Kahl, 1989; Needham, Teel, 1991).
Как и другие наземные членистоногие, клещи имеют отрицательный водный баланс. Они могут существовать только благодаря восполнению потерь воды за счет ее активного поглощения и снижения транспирационных потерь. Величина последних определяется разницей между содержанием водяных паров в атмосфере, выражаемой в единицах дефицита насыщения или процентах относительной влажности, и активностью водяных паров солевого раствора (гемолимфы) организма клеща. Активность водяных паров гемолимфы выражается в единицах осмотической концентрации, по точке замерзания, эквивалентному значению раствора NaCl или в виде доли от давления водяных паров над поверхностью чистой воды, принятого за единицу.
Осмотическая концентрация гемолимфы для немногих исследованных видов составляла у взрослых 190—380 мМ/л NaCl (Kaufman, Sauer, 1982), что эквивалентно 0.99 давления водяных паров при норме 100 %. В естественных микробиотопах клещей относительная влажность бывает, за немногими исключениями, значительно ниже, и градиент перемещения воды направлен из организма наружу в атмосферу. Даже в наиболее благоприятных для клещей местообитаниях они постоянно расходуют воду на испарение, и одними из главных проблем их существования являются сохранение воды в организме и возмещение ее потерь.
Потери воды складываются из ее испарения сквозь покровы и из трахейной системы, а также при экскреции и дефекации (рис. 72). У голодающих особей два последних компонента сведены до минимума и основные потери воды связаны с транспирацией. Дефекация происходит в первые дни после питания, а затем у личинок и нимф она прерывается и возобновляется уже после линьки. У голодающих особей дефекация также происходит со значительными перерывами. Главный продукт азотистого обмена клещей — гуанин обладает очень малой растворимостью в воде и скапливается в виде взвеси сферических конкреций в полости мальпигиевых сосудов и ректального пузыря (Hamdy, 1973). В ректальном пузыре экскреты смешиваются с поступающими в него из кишечника фекальными массами, и в этом же органе происходит интенсивная реабсорбция воды. Соотношение между экскретами и фекалиями в продуктах дефекации клещей на разных стадиях жизненного цикла меняется. В результате окраска фекалий также варьирует от почти белой из-за преобладания гуанина до черной или бурой при выведении гематина и других продуктов неполного переваривания крови. В еще большей степени меняется консистенция фекальных масс от почти жидкой во время питания, сразу после него и в первые дни по окончании линьки до твердых обезвоженных шариков у голодающих клещей.
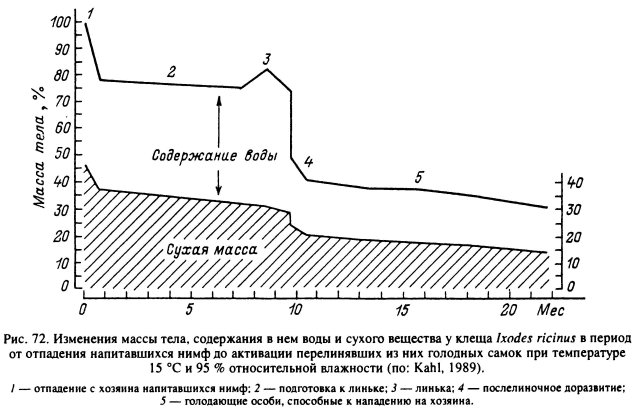
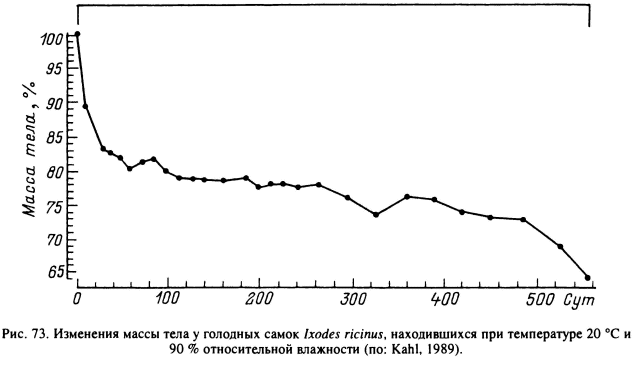
Помимо стадии питания, когда из организма выводится большое количество лишней воды в процессе обезвоживания поглощенной крови, значительное количество воды выводится из организма в процессе дефекации у свежеперелинявших особей на стадии послелиночного доразвития. Например, у взрослых A. hebraeum в первые 8 сут после линьки самки весят в среднем 44 мг и самцы — 29 мг, а через 4—6 нед их масса уменьшилась более чем на 20 %, главным образом за счет дефекации. В дальнейшем масса голодных особей, как и содержание воды в их организме, стабилизировалась. На протяжении 10 нед наблюдений при температуре 20 °С и 93 % влажности самки весили в среднем 32 мг, самцы — 23 мг, а содержание воды в их организме составляло соответственно 60.2 ± 0.7 и 60.3 ± 1,0 % (Rudolph, Knulle, 1978). Сходная динамика весовых изменений в течение более 500 сут после линьки отмечена и для самок I. ricinus (рис. 73) (Kahl, 1989).
По-видимому, у закончивших послелиночное доразвитие голодных клещей при относительной влажности от 85 % и выше содержание воды в организме относительно постоянно и составляет 60—70 %. В экспериментах с разными видами было установлено, что голодающие клещи могут терять за счет испарения до 20—40 % содержащейся в них воды, после чего наступает утрата подвижности и затем, при больших потерях, необратимые повреждения и гибель. У голодных самок Н. asiaticum смерть наступала после потери 50 % воды (Балашов, 1960), а у самок A. americanum — даже 70 % (Sigal, 1990).
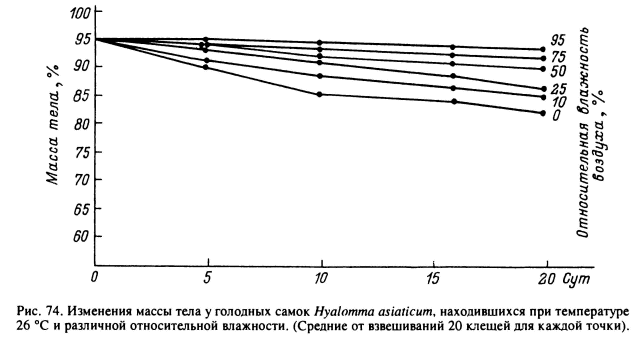
Многочисленными полевыми и лабораторными экспериментами было обнаружено, что устойчивость разных видов иксодид к высыханию значительно варьирует и в общем соответствует увлажненности их естественных местообитаний. Наименьшая устойчивость свойственна I. ricinus, голодные самки которого выживали при О % относительной влажности и температуре 25 °С не более 1—2 сут. Они теряли за этот период до 50 % исходного содержания воды в организме (Lees, 1946). У мезофильного A. americanum среднесуточные потери воды в аналогичных условиях достигали 7—10 % массы тела (Sauer, Hair, 1971), а у ксерофильных R. sanguineus и D. marginatus — только 2—3 % (Lees, 1946). У голодных имаго Н. schulzei, Н. anatolicum excavatum и R. bursa при 0 % влажности и температуре 25 °С среднесуточные потери массы составляли всего 1 % (Rudolph, Knulle, 1978). Наибольшая устойчивость к высыханию обнаружена у Н. asiaticum (Балашов, 1960). Голодные имаго этого вида безболезненно теряли до 30 % воды, содержащейся в их организме, и быстро восполняли эти потери после перенесения их в условия высокой влажности (>75 %). При температуре 26 °С и относительной влажности 10 % клещи за 1 сут теряли около 0.6 % массы тела и выживали до 1 мес и более (рис. 74).
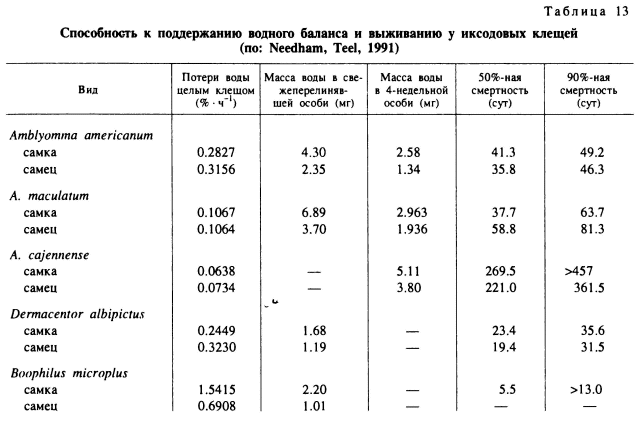
Связь между степенью устойчивости клещей к высыханию и аридностью или мезофильностью их местообитаний выявлена для 5 видов, живущих на юго-востоке США. При содержании клещей в эксикаторе (табл. 13) наименьшие потери воды организмом (% • ч-1) были у взрослых A. cajennense (0.0638—0.0734), отличающихся наибольшей ксерофильностью. Затем в убывающем порядке следовали А. maculatum, A. americanum, D. albipictus и В. annulatus (1.5415 у самок и 0.6908 у самцов) (Needham, Teel, 1991). Сходная связь между увлажненностью местообитаний и устойчивостью к высыханию обнаружена и для напитавшихся личинок 3 видов клещей, обитающих в Австралии. При высоком дефиците насыщения у личинок гигрофильных Н. longicornis и I. holocyclus масса тела уменьшалась быстрее, а сроки жизни были короче, чем у ксерофильного R. sanguineus (Heath, 1981).
Скорость транспирационных потерь воды определяется относительной влажностью воздуха, а более точно — дефицитом насыщения атмосферы водяными парами. Последний показатель непосредственно связывает комбинированное воздействие температуры и влажности на живой организм, но в экологической литературе он используется реже, чем более просто определяемая относительная влажность воздуха. Потери воды клещами растут с уменьшением относительной влажности воздуха и увеличением температуры, а сроки жизни при этом соответственно сокращаются.
Например, имаго Н. asiaticum при 26 °С выживали при относительной влажности от 0 до 25 % до 20 сут и более (срок наблюдений), а при 36 °С в 10 %-ной влажности — 10 сут и в 50—95 %-ной — 20 сут (срок наблюдений) (Балашов, 1960). У другого пустынного вида — Н. dromedarii при температуре 28 °С голодные самки теряли за 25 сут за счет испарения при 42 % относительной влажности 24.6 % своей исходной массы, при 60 % — 17.4 и при 75 % — 9.7 % и жили соответственно 83, 119 и 142 сут (Hafez et al., 1970). Голодные самки клеща R. evertsi mimeticus выживали в 50 %-ной относительной влажности при температуре 40 °С 30.5 сут, при 30 °С — 48.3 и при 10 °С — 100.7 сут, а при 15 % влажности соответственно 18.2, 25.9 и 69.Q сут (Kalvelage et al., 1987). Личинки В. microplus при температуре 22 °С выживали при 90 % относительной влажности 240 сут, а при 70 % — только 12 сут (Hitchcock, 1955).
Величина транспирационных потерь при прочих равных условиях зависит не только от видовой принадлежности, но и значительно различается для разных морфологических фаз и стадий развития одного вида. Наименьшей устойчивостью к высыханию обладают яйца, большей — личинки и нимфы и наибольшей — взрослые особи. Например, у Н. dromedarii при 42 % относительной влажности и 28 °С голодные личинки выживали 4—5 сут, нимфы — до 16, самки — до 83 и самцы — до 101 сут (Hafez et al., 1970).