Иксодовые клещи — паразиты и переносчики инфекций
Сохранение воды в организме
Мелкие размеры тела клещей и особенно их личинок обусловливают высокие значения отношения поверхности тела к его объему и соответственно высокие потери воды на испарение. В подобной ситуации важная роль принадлежит приспособлениям для снижения испарения и консервации воды в организме. Среди подобных адаптации первое место принадлежит водонепроницаемости кутикулы и наличию специализированного замыкательного механизма в стигмах. Потери воды уменьшаются также благодаря выведению конечных продуктов азотистого обмена в форме малорастворимого в воде гуанина и реабсорбции воды из экскретов и фекалий. Дефекация и экскреция приурочены к ограниченным периодам в жизненном цикле, когда в организме сохраняется запас избыточной воды после питания.
Кутикула, покрывающая поверхность всего тела клещей и выстилающая стенки трахей, служит барьером между внутренней средой организма и окружающей средой. Водонепроницаемость кутикулы относительна. Она значительно различается у разных видов по эффективности снижения водопотерь. Во всех случаях при уменьшении относительной влажности ниже определенного для каждого вида порогового значения — так называемой критической влажности клещи начинают интенсивно терять воду за счет испарения и масса их тела быстро уменьшается.
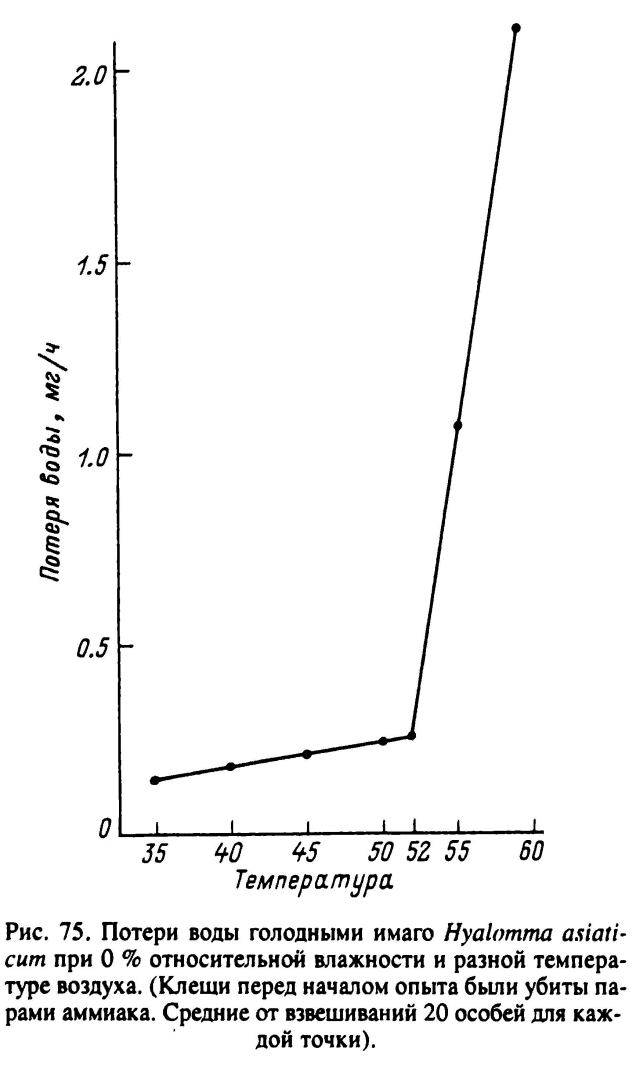
Все слои кутикулы обладают водонепроницаемостью, но главную роль в этом играет самый поверхностный восково-липоидный слой. Проницаемость кутикулы для воды увеличивается с повышением температуры, и по достижении определенных ее значений, при так называемой критической температуре кутикулы, происходит резкое увеличение скорости испарения воды клещом. Это явление объясняют нарушением молекулярной ориентации в восково-липоидном слое. По величинам критических температур выявлены четкие видовые различия, отражающие биотопическую приуроченность разных групп иксодид (Lees, 1947; Балашов, 1967; Knulle, Rudolph, 1982). Наиболее низкая критическая температура кутикулы — 32 °С обнаружена у гигрофильного I. ricinus, у обитателей более сухих биотопов D. andersoni, R. sanguineus и Н. anatolicum она близка к 44—45 °С и у наиболее ксерофильного H. asiaticum — 52 °С (рис. 75). Абсолютные значения критических температур кутикулы близки к верхним температурным границам жизни соответствующих видов клещей.
Расправление складок кутикулы у питающихся особей существенно увеличивает ее площадь и тем самым испаряющую поверхность. Тем не менее для многих видов, включая H. asiaticum (Балашов, 1989в) и A. americanum (Needham, Teel, 1991), напитавшиеся клещи устойчивее, чем голодные, к высыханию. Это обусловлено выделением больших количеств восково-липоидных веществ дермальными железами питающихся клещей, как показано для самок В. microplus. В результате у напитавшихся особей на поверхности кутикулы образуется новый защитный водонепроницаемый слой с хорошо выраженными гигрофильными свойствами (Knulle, Rudolph, 1982).
Существенные потери воды связаны с дыханием. Приспособлением для их снижения, по-видимому, является периодичность воздухообмена трахейной системы с внешней средой. Трахейная система открывается наружу на паре стигмальных пластинок через множество мелких отверстий, образующих аэропиль (табл. XII, 7). Под стигмальной пластинкой находится обширная воздушная полость, на границе которой располагается замыкательный аппарат, отделяющий полость от входа в главный трахейный ствол (табл. XII, 2). В нормальных условиях ритмика газообмена определяется чередованием периодов открывания и закрывания замыкательного аппарата. В экспериментах с содержанием клещей в атмосфере с высоким содержанием СО2 было показано, что трахейная система постоянно открыта и замыкательный аппарат не препятствует воздухообмену. В этих условиях у голодных взрослых A. variegatum при температуре 20 °С и 0 % относительной влажности потери воды возрастали в 17 раз. В состоянии покоя дыхальца этих клещей бывают открыты 1—2 раза в час по 4— 7 мин. Именно в эти короткие периоды и происходит достоверное уменьшение массы тела за счет транспирации. У двигающихся особей дыхальца бывают открыты до 15 раз в 1 ч и соответственно у них резко увеличены потери воды (Rudolph, Knulle 1979; Kniille, Rudolph, 1982). Сходный прерывистый тип газообмена обнаружен у голодных взрослых A. hebraeum. У неподвижных самок этого вида число выбросов СО2 в атмосферу составляло в среднем 1 за каждые 80 мин. У напитавшихся особей газообмен непрерывен и активная регуляция, по-видимому, отсутствует (Fielden et al., 1994). У личинок иксодид трахейная система отсутствует, дыхание осуществляется через покровы тела и изменения концентрации СО2 в атмосфере не влияют на» скорость потерь воды организмом.