Иксодовые клещи — паразиты и переносчики инфекций
Продолжительность питания
Для всех трех фаз развития иксодовых клещей характерно многодневное питание при однократном приеме пищи на каждой фазе. У большинства видов, изученных в этом отношении, питание личинок на млекопитающих и птицах продолжается 3—6 сут. У видов Amblyomma сроки питания всех фаз несколько дольше, чем у других родов: у нимф — 3—10, у самок — 6—12 сут (табл. 3). Самцы клещей приобретают способность к спариванию и осеменению самок спустя 3—6 сут после прикрепления. Однако после спаривания они не оставляют хозяина,но остаются присосавшимися к нему от нескольких недель до 2—3 мес и дольше.
Все это время они периодически поглощают свежую кровь и в поисках новых самок для спаривания могут менять места прикрепления.
Исключения составляют личинки и нимфы примитивнейшего подрода Alloceraea из рода Haemaphysails. У личинок Н. inermis при паразитировании на лабораторных грызунах питание занимает от 2.5—3.5 ч до 2 сут, у нимф — от 4 ч до 2 сут (Nosek, 1973; Успенская, 1987), тогда как самки и другие иксодиды напитываются за 6—7 сут. У Н. kitaokai личинки напитывались за время от 2 до 20 ч, нимфы — от 3.5 до 6 ч, а самки на крупном рогатом скоте — за 8—15 сут (Kitaoka, Morii, 1967).
У двух- и однохозяинных видов продолжительность собственно питания, если вычесть периоды подготовки к линьке, та же, что и у треххозяинных видов. Например, у однохозяинного В. microplus Личинки насыщаются в основной массе за 3 сут, но у части особей питание продолжается до 6 сут; нимфы питаются 4—7, а самки — 5—10 сут (Nunez et al., 1985) (табл. 5). У двуххозяинных клещей R. bursa (Балашов, 1967) и R. evertsi (Rechav et al., 1977) питание у личинок занимает 6—8 сут. У клеща Н. marginatum rufipes личинки питались на кролике 8—15, а нимфы — 6—15 сут (Knight et al., 1978) (табл. 4).
Средние сроки питания иксодид не являются репрезентативным видовым признаком этих паразитов. Даже в условиях эксперимента, при кормлении партий клещей одного календарного и физиологического возраста на лабораторных или сельскохозяйственных животных, период отпадения напитавшихся особей у личинок и нимф занимает 3—7 сут, а самок — 5—10 и более. Особенно сильно растянуты сроки отпадения напитавшихся нимф двуххозяинных видов и самок однохозяинных. Например, нимфы Н. marginatum, напитавшиеся на кроликах, отпадают в течение 10—25 сут, нимфы R. bursa — до 11 сут, а самки В. annulatus при питании на крупном рогатом скоте — не менее 2—3 нед (Балашов, 1967). К сожалению, причины значительных индивидуальных различий в сроках питания клещей исследованы недостаточно. Продолжительность питания клеща отражает суммарный результат взаимодействия многих факторов и в первую очередь зависит от физиологического состояния самого паразита, многих особенностей организма хозяина и прямого или опосредованного воздействия факторов окружающей среды. Вычленить роль каждого из этих факторов затруднительно даже в условиях эксперимента.
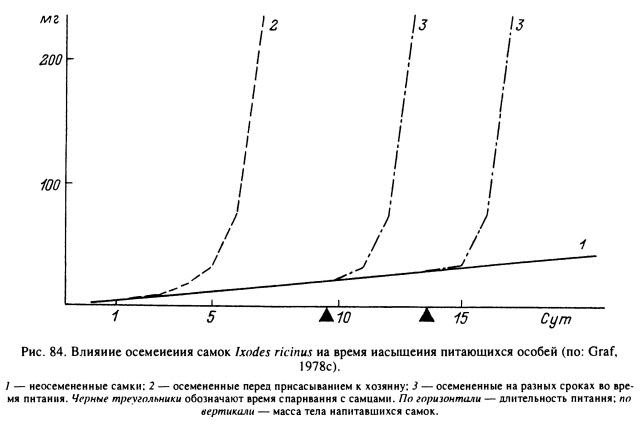
Значительное удлинение сроков питания самок может быть связано с задержкой копуляции питающихся особей. Для нормального насыщения самок рода Ixodes они должны быть оплодотворены еще голодными, или же спаривание происходит спустя 4—6 сут после начала питания (рис. 84). Неоплодотворенные самки Amblyomminae нормально питаются 5—6 сут, а затем их размеры и масса увеличиваются очень медленно (рис. 85). Такие особи могут оставаться прикрепленными на теле хозяина в течение нескольких недель, но не достигают нормального насыщения. Некоторые из них в конечном итоге отпадают недопитавшимися и даже откладывают небольшое количество неоплодотворенных яиц, а другие погибают, оставаясь прикрепившимися. В случае спаривания с самцами самки благополучно завершают питание в течение 1—2 сут и, достигнув нормальной массы, покидают хозяина. В естественных условиях на хозяине обычно самцов больше, чем самок, что увеличивает вероятность своевременного спаривания, и, следовательно, длительные задержки в отпадении редки (Балашов, 1967; Diehl et al., 1982).
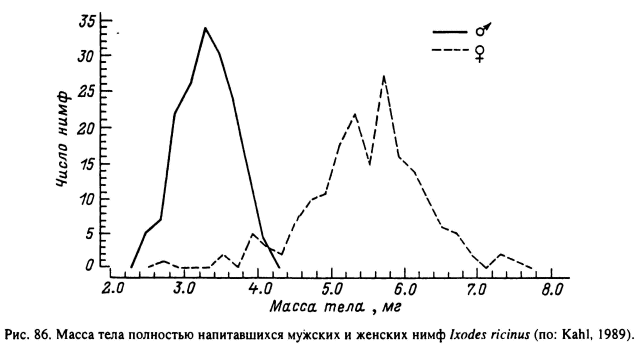
Для нескольких видов родов Rhipicephalus, Hyalomma и Amblyomma обнаружено, что сроки насыщения мужских нимф немного короче, чем женских (Feldman-Muhsam, Muhsam, 1966; Branagan, 1974; Norval et al., 1980). У I. persulcatus и
I. ricinus женские нимфы, которые достоверно отличаются от мужских большей массой тела (рис. 86), питаются на 8—9 ч дольше, чем мужские (Таежный клещ..., 1985; Kahl, 1989). Сроки паразитирования клещей могут удлиняться на 12—24 ч и более в связи с задержкой отпадения полностью напитавшихся особей. Напитавшийся клещ, по-видимому, определяет момент освобождения своих ротовых органов из кожи хозяина и отпадения получением определенных сигналов со стороны организма прокормителя или окружающей среды. Наиболее детально исследовано влияние светового режима и в меньшей степени температуры, которые служат пусковыми механизмами для рассмотренных в главе IV циркадных ритмов отпадения напитавшихся особей (табл. 7).
В лабораторных экспериментах с регулируемой продолжительностью темного и светлого периодов суток было установлено, что отпадение напитавшихся личинок и нимф A. hebraewn с кроликов происходит в самом конце светового или начале темного периода. Этот процесс регулируется эндогенным циркадным ритмом организма клеща, который имеет фотопериодическую настройку. Решающая роль принадлежит фотопериодическим условиям во время питания клеща, хотя для некоторых видов отмечено влияние фоторежима еще на стадии голодной особи (Rechav, 1978). При нарушениях нормального ритма освещения созданием полного затемнения или круглосуточного освещения, как было показано для Н. leporispalustris (George, 1971), эндогенный контроль отпадения клещей нарушается. Регуляция отпадения начинает осуществляться различными экзогенными факторами и в первую очередь физиологическими и биохимическими процессами в организме хозяина, обладающего собственным циркадным ритмом. Сходная ситуация была обнаружена у напитавшихся самок I. persulcatus (Балашов, 1954) при изменении времени выпаса крупного рогатого скота с дневных часов на ночные. В результате отпадение напитавшихся особей также переместилось на ночь, а поглощение последней, наибольшей по объему, порции крови — на день.
При паразитировании клещей одного вида на интактных особях разных видов их природных хозяев, за исключением рептилий, и на лабораторных животных сроки питания могут отличаться. Чаще подобные различия недостоверны в связи с очень большим размахом индивидуальной изменчивости. По, нашим наблюдениям (Балашов, 19586), в опытах с кормлением личинок и нимф I. ricinus, D. reticulatus, D. turanicus, D. asiaticum на кроликах, морских свинках, белых мышах и сирийских хомяках, а самок — на кроликах, морских свинках, овцах и крупном рогатом скоте при прочих равных условиях средние сроки от посадки клещей на хозяина до отпадения напитавшихся особей варьировали в пределах 1—3 сут. Кормление личинок I. persulcatus на обыкновенных полевках (Microtus arvalis), рыжих полевках (Cletrhrionomys glareolus), лесных мышах (Apodemus sylvaticus), дроздах-рябинниках (Turdus pilaris), белых мышах, морских свинках, кроликах не выявило достоверных различий в сроках насыщения. Аналогичные результаты были получены в опытах с личинками D. asiaticum и D. marginatus (Балашов, 19936). В опытах Н. Н. Лебедевой (Лебедева, 1980, 1981; Лебедева, Платова, 1984) личинки I. persulcatus питались на лесных мышах немного быстрее (3.6 сут), чем на трех видах полевок (4.2 сут), а личинки D. reticulatus также быстрее насыщались на лесных мышах и степных пеструшках (Lagurus lagurus) и медленнее на 2 видах полевок. Однако эти различия не были статистически достоверными.
В отличие от млекопитающих и птиц сроки насыщения клещей на рептилиях значительно продолжительнее и более вариабельны. Например, у А. marmoreum при температуре 26 °С питание личинок на черепахе Geochelone pardalis продолжалось в среднем 30 сут, нимф — 51 и самок — 60 сут (Norval, 1975b). У I. persulcatus в природных условиях личинки питались на ящерицах 13—17 сут (Халанский, Положихина, 1962). В эксперименте установлено, что сроки насыщения клещей на рептилиях зависят от температуры окружающей среды. Так, при температуре 20 °С личинки I. ricinus питались на прытких ящерицах (Lacerta agilis) 8—12 и нимфы — 11—15 сут, при 25 °С — соответственно 6—11 и 6— 10 сут, а при 30 °С — 3—4 и 4—7 сут. В аналогичных условиях сроки насыщения личинок и нимф Н, sulcata при 20 °С были в 2—3 раза продолжительнее, чем при 30 °С (Балашов, 1967). При кормлении личинок и нимф Н. aegyptium на черепахе Testudo graeca и других рептилиях при температурах от 8 до 37 °С ниже 20 °С полное насыщение клещей невозможно, хотя прикрепление паразитов к хозяину происходило при температуре более 10 °С. В диапазоне от 20 до 30 °С понижение температуры на 1 °С вызывало пропорциональное удлинение сроков насыщения личинок на 1.5 сут и нимф — на 3.25 сут. Соответственно при 20 °С питание продолжалось 3—9 и 10—15 сут, а при 30 °С — 19—34 и 33—65 сут (Sweatman, 1970).
При питании иксодовых клещей на теплокровных животных изменения температуры окружающей среды мало влияют на сроки их насыщения. Температура тела у большинства млекопитающих обладает высоким постоянством, хотя и может быть различной на разных частях тела, а на поверхности кожи эти колебания достигают максимальных значений. Тем не менее размах колебаний температуры в местообитаниях клещей на теле млекопитающих и птиц несравнимо меньше, чем у рептилий. У грызунов и других животных, обитающих в норах или ведущих ночной образ жизни, подобные суточные колебания выражены еще слабее. В результате на теплокровных животных клещи находятся при температуре от 25 до 35 °С, и подобный тепловой режим характерен для многих, даже неродственных видов хозяев. Поэтому сроки питания клещей на разных видах хозяев из млекопитающих и птиц могут существенно не отличаться, а выявленные различия могут определяться не температурой, а иными факторами (Балашов, 1982). Исключение составляют некоторые виды рукокрылых, насекомоядных и грызунов, впадающих в зимнюю спячку, у которых температура тела может значительно понижаться и приближаться к таковой в норе или в другом убежище. Этим фактором можно, в частности, объяснить удлинение сроков насыщения I. ricinus на ежах, находящихся в состоянии зимней спячки (Carrick, Bullough, 1940).
Во многих случаях удлинение сроков питания бывает вызвано прикреплением клещей к неподходящим местам на теле хозяина с недостаточным кровоснабжением, обильными выделениями жиропота, ороговением. Из всех исследованных факторов организма хозяина наибольшее влияние на сроки питания и другие показатели этого процесса оказывает иммунное состояние его организма (см. главу VII).