Иксодовые клещи — паразиты и переносчики инфекций
Факторы, влияющие на противоклещевую резистентность
Приобретенная противоклещевая резистентность в большинстве случаев не абсолютная и препятствует развитию на хозяине лишь части напавших на него паразитов. Ее количественные показатели (процент погибших особей, масса тела и др.) могут значительно варьировать не только среди животных одной породы и в одном стаде, но и у одних и тех же особей в разные сезоны и годы, в связи с климатическими колебаниями и под влиянием других, часто трудно оцениваемых факторов внешней среды. В еще большей степени устойчивость может меняться в зависимости от физиологического состояния животных и особенно их возраста и пола. Поэтому во многих случаях трудно вычленить воздействие на питание и последующее развитие клещей собственно иммунных механизмов от не связанных с ними факторов окружающей среды и организма хозяина.
Влияние различных факторов на противоклещевую устойчивость наиболее детально исследовано на примере крупного рогатого скота и однохозяинного клеща В. microplus. Разные породы и даже отдельные животные одной породы могут существенно отличаться по способности к развитию приобретенной устойчивости. Породы азиатского зебувидного скота оказались значительно устойчивее европейских пород. У несенсибилизированных животных, безотносительно к их породе, при первичной посадке на них голодных личинок стадии напитавшихся самок достигало около 20 % от выпущенных особей. Через месяц после развития иммунитета при повторных кормлениях личинок на зебувидном скоте этот же показатель был не более 1 %, а на европейском скоте — 5—10 %. Способность к развитию противоклещевой устойчивости против В. microplus наследуется, и у гибридов она положительно коррелирует с долей крови от зебувидных родителей (Wagland, 1975, 1978а, 1978b; Utech et al., 1978; Sutherst et al., 1988b; Miranpuri, 1989).
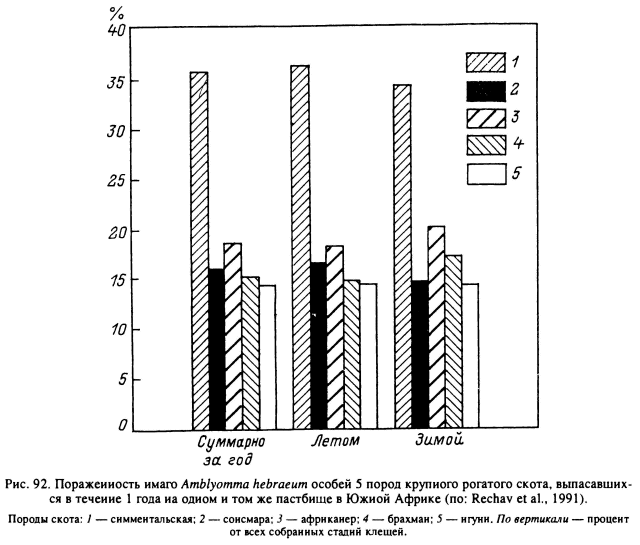
Генетическая детерминированность устойчивости крупного рогатого скота против клещей выявлена и для других видов иксодид (рис. 92). Например, в Восточной Африке при естественном выпасе на одних пастбищах животных разного происхождения на гибридах Bos indicus x В. taurus паразитировало в среднем в 4—5 раз больше имаго H. anatolicum (70.5 и 17 особей), чем на местных породах, близких к В. indicus, г масса напитавшихся самок составляла соответственно 422 и 375 мг (Latif, 1984b). В эксперименте с нимфами того же вида на резистентном гибридном скоте заканчивало питание только 10 % от выпущенных клещей, и средняя масса напитавшихся нимф составила 11.3 мг. На 2 группах чувствительных животных европейского происхождения напитывались 20 и 45 % нимф, масса тела которых была в среднем 15.8 и 13.6 мг (Latif, 1984a). Наследственная способность к развитию противоклещевой устойчивости у крупного рогатого скота варьировала в зависимости от пола и возраста таким образом, что самцы обычно поражены сильнее самок, а молодые животные меньше взрослых (Tatchell, 1986; Sutherst et al., 1988b). На развитие резистентности влияли упитанность животных, состояние беременности или лактации, а также такие факторы окружающей среды, как сезонные изменения длины дня и выпадение осадков (Sutherst et al., 1988a; Bourne et al., 1988).
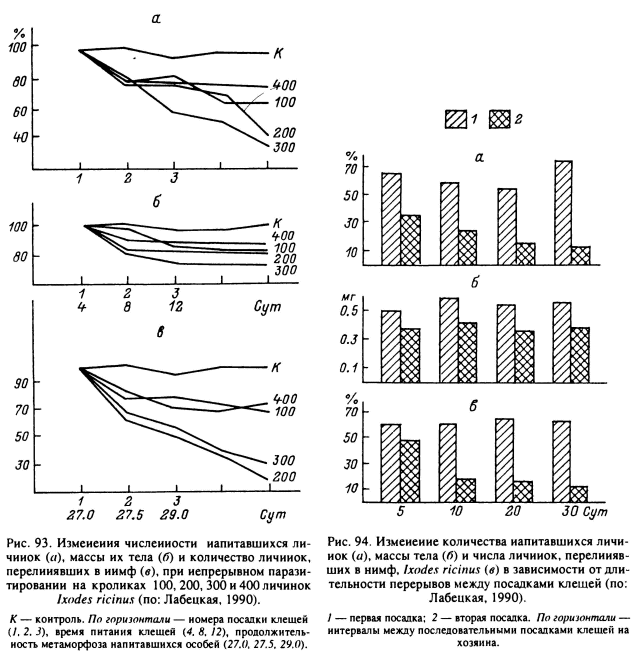
Уровень противоклещевой резистентности может определяться количеством одновременно питающихся клещей и промежутками времени между их повторными паразитированиями. А. Г. Лабецкой (1990) было показано, что при непрерывном паразитировании на кролике 200—300 личинок или 25 нимф I. ricinus (рис. 93), а на рыжей полевке — 5 личинок через 13—19 сут от начала питания (после 3-или 4-кратного срока насыщения) хозяева сенсибилизировались. Это ухудшало условия питания последующих партий паразитов. Непрерывное паразитирование больших количеств клещей, а именно 400 личинок, 100 нимф или 5—10 самок на кролике, а также 20—50 личинок на рыжей полевке, напротив, не вызывало развития резистентности. В этих же экспериментах показано, что животные приобретали устойчивость при перерывах между кормлениями клещей в 10 сут и более. При снижении интервала до 5 сут уровень резистентности был значительно ниже (рис. 94).
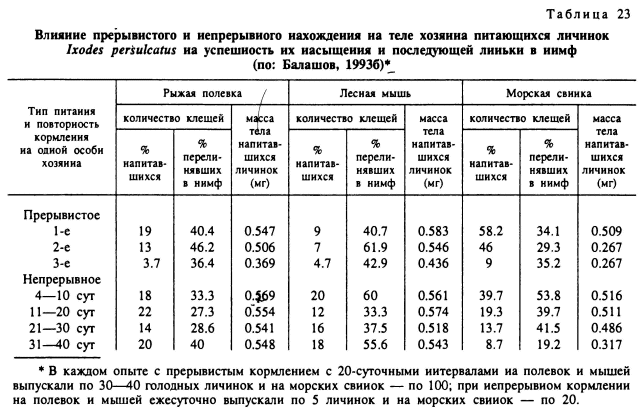
В моих опытах (Балашов, 19936) у лесных мышей и рыжих полевок при непрерывном паразитировании на них личинок I. persulcatus, а у краснохвостых песчанок и ушастых ежей — личинок Н. asiaticum противоклещевая резистентность не развивалась или была очень слабой (табл. 23). Процент напитавшихся личинок, масса их тела и способность к линьке в нимф достоверно не различались для особей, питавшихся в первые и последние 10 сут 40-суточного эксперимента. Повторные кормления личинок этих же клещей и на тех же видах хозяевах, но с разрывами в 20 сут между окончанием одного и началом следующего питания стимулировали выработку достаточно высокого уровня резистентности.
В условиях эксперимента при последовательном кормлении на овцах партий имаго Н. asiaticum в зависимости от частоты повторных посадок и количества питающихся клещей у разных животных развивались как состояния противоклещевой резистентное, так и десенсибилизации, не препятствующей нормальному насыщению паразитов (Бердыев, Худайназарова, 1976). Отсутствие или слабая резистентность в ответ на многократные повторные кормления отмечены для имаго A. hebraeum на крупном рогатом скоте и овцах (Norval et al., 1988a) и для нимф и личинок этого вида на овцах и кроликах (Norval, 1978).
Возникновение резистентности как защитной реакции на питание клещей было обнаружено и исследовано главным образом у лабораторных животных (морские свинки, кролики, белые мыши) и крупного рогатого скота. Повторные питания клещей на этих хозяевах в большинстве опытов были менее успешными, чем первичные на несенсибилизированных особях. Напротив, у некоторых видов диких животных — естественных хозяев клещей питание специфичных для них паразитов не вело к развитию резистентности. В ряде случаев, при кормлении одного вида клеща на хозяевах разных видов, резистентность развивалась у лабораторных животных, но была слабо выражена у диких животных — природных хозяев этих клещей.
Впервые это явление было обнаружено при лабораторном кормлении личинок D. variabilis на его главном прокормителе в Северной Америке — грызуне Peromyscus maniculatus. Для возникновения умеренной резистентности у этого вида требовалось 2—3 повторных кормления партиями из 400 личинок, тогда как для выработки почти полной устойчивости у морских свинок было достаточно однократного кормления 100 личинок (Trager, 1939).
В опытах с кормлением I. trianguliceps на одном из его природных прокормителей — лесных мышах Apodemus silvaticus повторные кормления по 30—50 личинок или 8 нимф на одном зверьке не привели к развитию у них резистентности (табл. 24). Процент прикрепившихся и закончивших питание клещей и масса тела по отпадении после 2—4-го кормления существенно не отличались от тех же показателей после 1-го кормления. Напротив, при кормлении того же вида на белых мышах у последних развивалась противоклещевая резистентность (Randolph, 1979). Сходно вели себя и главные прокормители I. scapularis — полевки Microtus pensylvanicus. 3 повторных кормления партий по 100 личинок на полевках не вызвали достоверного отторжения или гибели клещей, тогда как при повторном питании на морских свинках развитие заканчивало не более 5 % особей (Davidar et al., 1989; Riberio, 1989).

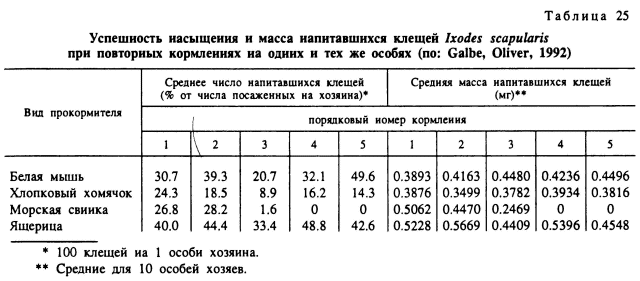
При 5 последовательных кормлениях личинок этого вида на хлопковых крысах (Peromyscus gossipinus), белых мышах, морских свинках и ящерицах (Eumeces laticeps) высокая резистентность к повторным питаниям также была свойственна только морским свинкам, но полностью отсутствовала у ящериц и была слабо выражена у 2 других видов грызунов (табл. 25) (Galbe, Oliver, 1992). Аналогичные результаты были получены при 3-кратном кормлении личинок A. marmoreum и A. hebraeum на морских свинках, цесарках и черепахах, когда резистентность не возникала только у одного из главных хозяев обоих видов в природе — цесарок (Fielden et al., 1992). Резистентность не возникала после повторных кормлений личинок I. persulcatus на их главных хозяевах — серой и рыжей полевках (Лебедева, 1980). Отсутствие резистентности к этому виду клещей было отмечено и в наших опытах с кормлением личинок на рыжих полевках, лесных мышах, но только в случае непрерывного паразитирования. В то же время она возникала при кормлении партий личинок с 20-суточными перерывами (Балашов, 19936). Подобным же образом реагировали рыжие полевки на повторное или непрерывное паразитирование на них личинок и нимф I. ricinus (Лабецкая, 1990).
Различный уровень иммунитета был выявлен у диких животных в их естественных местообитаниях. В частности, у отловленных в природе полевок-экономок, красных и рыжих полевок, лесных и желтогорлых мышей процент напитавшихся на них в опыте личинок I. persulcatus был достоверно ниже при кормлении на грызунах из стаций с высокой численностью клещей и выше — из стаций с низкой численностью (Никитина, Аристова, 1964).
Существование положительной корреляции противоклещевого иммунитета с интенсивностью кожной реакции у позвоночных позволило оценить уровни резистентности в природных популяциях прокормителей. Так, в Замбии на пастбищах с высокой численностью A. variegatum и R. appendiculatus у особей крупного рогатого скота с минимальным количеством питающихся на них имаго (18 и 60 клещей соответственно) кожная реакция была наиболее интенсивной, а у максимально заклещевленных животных (1072 и 260) она практически не обнаруживалась (Jongejan et al., 1989). Подобная же отрицательная корреляция существует между числом клещей Н. leporispalustris, паразитирующих на флоридском кролике, и интенсивностью кожной реакции (McGowan et al., 1982).
Отсутствие резистентности после повторных питаний клещей на их естественных хозяевах Рибейро (Riberio, 1989) объясняет подавлением иммунных ответов на антигены определенными компонентами слюны этих паразитов. Реакция подавления резистентности видоспецифична и, по-видимому, возникла как взаимоадаптация партнеров по паразитарной системе. Ее проявления зависят от численности питающихся клещей, временных интервалов между повторными нападениями и других, трудно учитываемых факторов. Несмотря на отдельные исключения, эта гипотеза устраняет противоречия между высоким уровнем противоклещевого иммунитета у лабораторных и сельскохозяйственных животных и возможностью непрерывного паразитирования в природе в течение нескольких месяцев сменяющих друг друга клещей одного вида на одной особи хозяина.
В природе, и особенно в тропиках, на одной особи хозяина, одновременно или последовательно сменяя друг друга, могут паразитировать несколько видов клещей (рис. 95). К сожалению, степень видоспецифичности противоклещевой резистентности, возможности существования перекрестного иммунитета и иммунного взаимодействия разных видов клещей во время их питания на одном хозяине изучены очень слабо.
Противоклещевой иммунитет, как можно судить по многочисленным исследованиям, в значительной степени видоспецифичен. Данные Трегера (Trager, 1939) о возникновении у морских свинок перекрестного иммунитета между личинками D. variabilis, D. andersoni и даже Н. leporispalustris не получили полного подтверждения в более поздних работах. Оказалось, что между близкими видами одного рода — D. andersoni и D. variabilis у морских свинок действительно возникает перекрестный иммунитет. Он проявляется в снижении числа напитавшихся личинок «чужого» вида и массы их тела, но уровень его несколько ниже, чем по отношению к особям «своего» вида. В то же время между видами рода Dermacentor и представителями других родов — A. americanum или I. scapularis перекрестный иммунитет не возникал или был очень слабым (McTier et al., 1981). Отсутствовал перекрестный иммунитет между личинками R. appendiculatus, R. evertsi, A. hebraeum, A. variegatum и I. ricinus при кормлении их на морских свинках и кроликах (Rechav et al., 1989).
У крупного рогатого скота перекрестный иммунитет к близким видам — R. appendiculatus и R. pulchellus, по результатам анализа сроков питания нимф, массы тела напитавшихся особей и интенсивности кожной реакции к антигенам слюны «чужого» вида клещей, был слабым (DeCastro et al., 1989). Не был выявлен перекрестный иммунитет и у крупного рогатого скота, подвергавшегося на пастбище нападению клещей В. microplus и H. anatolicum (Miranpuri, 1989). Методами иммуноэлектрофореза в слюне нескольких видов иксодид были выявлены как видоспецифичные, так и групповые антигены. Некоторые из последних, по-видимому, ответственны за развитие перекрестного иммунитета, но его уровень значительно ниже, чем видоспецифичного (Almeida et ah, 1990; Jaworski et al., 1992).